- 1 起源与进化
- ▪ 起源
- ▪ 演化
- ▪ 藻类载色体及色素类型演化
- ▪ 形态演化
- 2 应用与研究历史
- ▪ 应用历史
- ▪ 研究历史
- ▪ 藻类体细胞育苗
- ▪ 藻类单倍体育种
- ▪ 耐高温性状选育
- 3 分类
- ▪ 分类系统变动
- ▪ 分类体系
- ▪ 国际分类体系
- ▪ 中国分类体系
- 4 代表类群及特征
- ▪ 褐藻门
- ▪ 海带目代表藻种
- ▪ 海带
- ▪ 裙带菜
- ▪ 墨角菜目代表藻种
- ▪ 红藻门
- ▪ 红毛菜目代表藻种
- ▪ 红盾藻目代表藻种
- ▪ 蓝藻门
- ▪ 念珠藻目代表藻种
- ▪ 色珠藻目代表藻种
- ▪ 其他藻门
- ▪ 绿藻门
- ▪ 甲藻门
- ▪ 金藻门
- ▪ 硅藻门
- 5 分布范围
- 6 生长发育
- ▪ 生长规律
藻类(Algae)是一类叶状植物的总称,它们没有真正的根、茎、叶分化,是以叶绿素a作为光合作用的主要色素,且在繁殖细胞周围缺乏不育的细胞包被物。藻类广泛存在于陆地和水体环境中,主要生活在水中,是地球上最常见的生物之一。目前已知的藻类种类已超过3万种。
起源与进化
编辑起源
藻类生物是非常独特的生物类群,其涵盖了原核生物和真核生物;包含了自养、兼养和异养几种不同的生活方式;囊括细菌类、植物类和动物类不同物种。因此,藻类的发生是具有多个层次的,发生于不同生物进化阶段;同时,不同藻类类群的起源是多元起源的,不具有单系起源的始祖。
对最古老形态化石的仔细研究表明,藻类的起源可追溯到33-35亿年前,最早的原核藻类蓝细菌(蓝藻)是地球生态系统自由氧出现的最初和最主要贡献者,并为需氧真核生物的进化提供了最主要的动力,而后其通过初级内共生事件在真核藻类的进化中亦发挥了重要作用。伴随着原核生物向真核生物的整体进化,出现了具有原始质体(蓝小体)的灰色藻门藻类,这也被认为是最早出现的真核藻类生物类群。大量数据表明,绿藻和红藻质体通过真核宿主和内共生体之间的高阶内共生合并,在真核生物树中广泛传播。由此可见,细菌在藻类起源中的作用是毋庸置疑的。
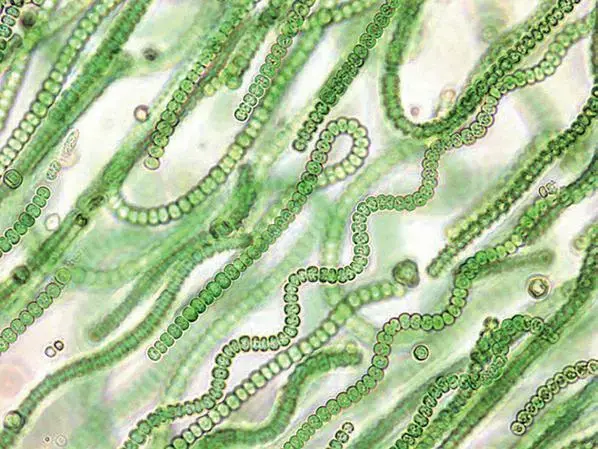

演化
藻类载色体及色素类型演化
除原核藻类外,所有真核藻类都有载色体。不同藻类植物所含的色素也是多种多样的,但归纳起有以下几大类:叶绿素类、类胡萝卜素(胡萝卜素和叶黄素)、藻胆素。根据藻体细胞载色体结构、所含光合色素及光合类型的不同,通常将藻类分为3条进化支系。最早形成的是原核蓝藻,大约比蓝藻晚了20亿年才出现真核红藻,形成了以藻胆素为光系统Ⅱ的主要集光色素这一演化路线,其次是含叶绿素a、叶绿素c的第二支系真核藻类的出现,最后才是第三支系,即叶绿素a、叶绿素b的真核藻类的产生。
形态演化
藻类是原始的低等植物,其个体通常称为叶状体。不同藻类的形态各异,有简单的单细胞藻体,有出现类似根、茎、叶结构分化的复杂、多细胞藻体。藻类形态的演化是一个由简单到复杂、从低级到高级的发展过程。典型的藻类形态体制有单细胞类型、群体类型、丝状体类型、膜状体类型和枝叶状体类型五种:
单细胞类型:原始、低级种类,藻体由单个细胞构成,例如原核的原绿藻 (Prochloyorsp.)和膨胀篮球藻(Chroococcus turgidus),以及真核的盐藻(Dunaliellasp.)等;
群体类型:由细胞直接相连或被胶质包埋而成,可分为能游动、不能游动和定形、不定形等类型;
丝状体类型:可分为:单列丝状体、 分枝丝状体和异丝体等类型。单列丝状体是由原始细胞向一个方向分裂,分裂后的细胞相互连接成一行的简单丝状体。分枝丝状体是单列丝状体内某些细胞向丝状体侧面单向分裂而成。异丝体亦是一种分枝丝状体,所不同的是由原始细胞分化为匍匐和直立的两部分,匍匐丝体具有固定直立丝体的作用;
膜状体类型:有真膜状体和假膜状体,真膜状体是由原始细胞向两三个方向分裂而形成的膜状结构,假膜状体是多个细胞或丝体被胶质包埋而成;
枝叶状体类型:最为复杂的类型,外形上有“根、茎、叶”的分化,内部有组织分化。
应用与研究历史
编辑应用历史
经考古发现,在14000年前,人类便收集藻类作为食物和药物,其中以海藻为主。海藻因其独特的风味和丰富的营养物质,在亚洲国家中广泛使用,自公元前500年以来中国便有着海藻消费记录;而在200多年前,海藻便在福建省平潭县有栽培养殖记录,在20世纪后,随着海藻养殖系统的成功建立和不断完善,中国已成为世界最大的海藻生产国。
在欧美国家,海藻是挪威、冰岛、葡萄牙、西班牙和爱尔兰等欧洲沿海国家的重要食物和肥料来源之一;在20世纪上半叶,欧美国家重点发展了海藻的工业应用领域,如提取用于食品加工的藻胶体;在20世纪中叶,由于第二次世界大战的影响,对从琼脂(属于藻胶体的一种)中生产的青霉素需求激增,使得对藻胶体产业的发展更为重视,而在20世纪末,葡萄牙成为了世界第五大琼脂的生产国。
研究历史
藻类体细胞育苗
关于藻类体细胞育苗研究主要包含藻类体细胞分化发育以及它们分化发育与内外界条件的关系两个方面。在了解其细胞分化发育的基础上,对温度、光照等外部条件进行研究,能更大程度地促进藻类细胞育苗的发展。1978年,美国加州大学N. 萨加等最早用液体培养法获得狭叶海带的感伤组织,并从中分离出单个细胞,且再生出孢子体。20世纪80年代以后,细胞培养进展迅速。1981年,中国学者赵焕登和张学成用研磨法;1983年,中国学者卢澄清用腐烂法:1984年,萨加用酶法分离到条斑紫菜单细胞并都再生成株。
藻类单倍体育种
海带、紫菜、裙带菜等具有异形世代生活史交替的大型海藻,可以通过藻类培养技术,获得纯合的单倍体生殖细胞,进而可按照有性生殖或者孤雌生殖模式,发育为纯合的二倍体植株。这种利用组织培养技术诱导藻类产生单倍体植株,再通过某种手段使染色体组加倍,从而使藻类恢复正常染鱼体数目的方法叫做藻类单倍体育种。利用海带雌性孢子体可以获得大量遗传稳定的磁性生殖细胞,可以进一步作为杂交育种的母本。19世纪末期,中国最早开展了海带单倍体育种工作,利用磁性孢子体产生的孢子单独买苗,与正常有性生殖的海带采苗的孢子放置于同一育苗系统中进行有性繁殖,培育出单倍体育种的子代,经过4代连续单棵自交选育,培育出海带“单海1号"品种。
耐高温性状选育
通过对经过物理、化学诱变处理或者未经过处理的群体,在高于其生物学生长发育适宜温度下进行培养,选择正常成活的个体作为亲本,自交培育子代。在选育过程中,通常结合看高产性状进行选育。海带是最早进行耐高温性状选育的藻类。20世纪60年代,中国藻类育种学家在配子体世代进行高温(18C和20°C)筛选,自交培育孢子体。在海上栽培养殖期间,挑选叶片长、自然脱落最轻的成熟海带作为亲本,采苗后在配子体阶段继续高温筛选,最终获得了海带“海青1号”宽叶品种和海带“海青2号”长叶品种。

紫菜育苗
分类
编辑分类系统变动
中国是记述海洋藻类最早的国家。公元前2至前1世纪的《尔雅》就已有关于海藻的记载,其后的历代本草和地方志中有更多的记述。1754年,分类学家林奈(Linnaeus)在《植物种志》中将植物界分为25个纲,并将藻类作为隐花植物纲四大类群之一;学者恩格勒(Engler,1844—1930)和勃兰特(Prantl,1849—1893)在《植物自然分科志(DieNaturlichen Pflanzenfamilien)》(1887—1915)提出的分类系统中,将藻类和真菌作为藻菌植物门(Thallophyta)独立的纲,首次确认了分类学地位。
1820和1824年,年瑞典海藻分类学者阿加德(Agardh)首次将红藻和褐藻予以区分,并确定了藻类的6个目。学者哈维(Harvey)于1836年明确提出以体色进行藻类类群的划分,设立了3个亚纲,绿色藻类(Chlorospermae,包括了蓝细菌和黄藻Xanthophycans),黑色藻类(Melanospermae)和红色藻类(Rhodospermae)。尽管当时恩格勒分类系统中的藻菌植物门(Thallophyta)中已设立了蓝细菌科(Cyanophyceae)、红藻科(Rhodophyceae)、 绿藻科(Chlorophyceae)和褐藻科(Pheophyceae),同时也包括了真菌。但作为“门”这一高级分类阶元(Phytoflagellates,植鞭毛虫类),则是由安德里歇(Endlicher)在1836年根据众多原生动物学家的提议设立的。
19世纪70年代中期,藻类学家们已对藻类的大部分属进行了描述,开展了部分种类的生活史研究,并对藻类分类系统不断地进行修正。其中,奥尔特曼斯(Oltmanns)在1922—1923年发表《藻类的形态与生物学(Morphologie und Biologie der Algen)》一书中,较为系统地总结藻类形态和生理、生态的研究成果。1935年,弗里奇(Fritsch)在《藻类的构造与生殖(The Structure and Reproduction of the Algae)》中,建立了较为完整的藻类分类依据体系。

红藻
分类体系
世界各国藻类学家对藻类的分类地位和系统演化尚未达成共识。
国际分类体系
国际上最早的藻类分类体系记录是学者福迪(Fott)在《藻类学(Algenkunde)》第二版中,根据学者Pascher(1931)的观点把藻类分为6个类群:蓝藻门(Cyanophyta)、杂色藻门(Chromophyta)(包括了金藻纲(Chrysophyceae)、黄藻纲(Xanthophyceae)、硅藻纲(Bacillariophyceae)、褐藻纲(Phaeophyceae)和甲藻纲(Dinophyceae))、红藻门(Rhodophyta)、绿藻门(Chlorophyta)等4个门;并把裸藻纲(Euglenophyceae)、隐藻纲(Cryptophyceae)、绿胞藻纲(Chloromonadophyceae)列为分类位置未确定的鞭毛类,把原胞藻目Protomonadales列为未确定分类位置的无色鞭毛类;学者Dawes在所著《海洋植物学(MarineBotany)》中,把海藻类分为8个门:蓝藻门(Cyanophyta)、绿藻门(Chlorophyta)、棕色藻门(Ochrophyta)、红藻门(Rhodophyta)、金藻门(Chrysophyta)、隐藻门(Cryptophyta)、裸藻门(Euglenophyta)、甲藻门(Pyrrophyta)。
最近的(2008年)藻类分类学是罗伯特·爱德华·李(Robert Edward Lee)在论著《藻类学(Phycology)》第四版中根据藻类质体内共生学说以及藻类质体内质体膜的层数,将藻类分为原核的蓝细菌门(Cyanophyta)以及真核藻类,真核藻类包括了灰色藻门(Glaucophyta)、红藻门(Rhodophyta)、绿藻门(Chlorophyta),质体单层内质体膜的裸藻门(Euglenophyta)、甲藻门(Dinophyta=Pyrrophyta)、顶复门(Apicomplexa),质体单层双质体膜的隐藻门(Cryptophyta)、异鞭藻门(Heterokontophyta)和普林藻门(Prymnesiophyta)共9门。
中国分类体系
在中国,20世纪80年代,曾呈奎等认为原绿藻(Prochlorondidemni)是光合生物进化独立分支,提出了藻类光合进化的观点,并以此把藻类分为12个门:蓝藻门(Cyanophyta)、红藻门(Rhodophyta)、隐藻门(Cryptophyta)、黄藻门(Xanthophyta)、金藻门(Chrysophyta)、甲藻门(Dinophyta=Pyrrophyta)、硅藻门(Bacillariophyta)、褐藻门(Phaeophyta)棕色藻门(Ochrophyta)原绿藻门(Chloroxybacteriaphyta)、裸藻门(Euglenophyta)、绿藻门(Chlorophyta)、轮藻门(Charophyta)。2005和2014年钱树本等编著的《海藻学》中也支持了曾呈奎的分类观点;2006年,胡鸿钧和魏印心编著的《中国淡水藻类一系统、分类及生态》中将藻类分为13个门,认为轮藻门(Charophyta)应被替换为灰色藻门(Glaucophyta)和定鞭藻门(Haptophyta)。由中国科学院中国孢子植物志编辑委员会编写的《海藻志》中,基本沿袭了曾呈奎的分类观点,已出版的门类包括了9个门:蓝藻门(Cyanophyta)、红藻门(Rhodophyta)、金藻门(Chrysophyta)、黄藻门(Xanthophyta)、硅藻门(Bacillariophyta)、褐藻门(Phaeophyta=棕色藻门Ochrophyta)、隐藻门(Cryptophyta)、甲藻门(Dinophyta=Pyrrophyta)和绿藻门(Chlorophytao)。
代表类群及特征
编辑藻类中褐藻门、蓝藻门、红藻门中种类较多,本章节着重介绍以上几门。
褐藻门
褐藻门(Phaeophyta)藻体为多细胞的异丝体、假膜体、膜状体,高级类群具有类似根、茎、叶等器官的分化,内部构造出现表皮、皮层和髓部等组织。细胞(除游动生殖细胞外)具有明显的细胞壁。色素体多为小盘状,亦有呈螺旋带状或分枝带状等。同化产物主要为褐藻淀粉和甘露醇等。生长方式为散生长、居间生长、毛基生长、顶端生长和边缘生长等。褐藻具有营养繁殖、无性繁殖和有性繁殖3种繁殖方式。褐藻门植物差异较大,小的仅几毫米,大的可达60米,甚至超过100米。褐藻广泛分布于全球海洋,淡水种类较少。根据经典形态学特征,本门被分为1纲16目(其他分类系统中被划分为21目),世界上有340余属2000余种,绝大部分生长在海洋里;中国有13目29科76属380余种。
海带目代表藻种
海带目(Laminariales)是指柄与叶片之间的分生组织通过居间生长形成二倍体的叶状体;其生殖方式为卵配。海带目的代表藻种有海带(Laminaria japonica Aresch)和裙带菜(Undaria pinnatifida Suringar)。
海带
海带的植物体为孢子体,较大,呈褐色,长2-3米,由上部扁平的带片、下部的带柄和基部分枝状的假根组成。带片最外层的表皮及皮下的皮层细胞中含有色素,如胡萝卜素、叶绿素和墨角藻黄素;带片的中央是由丝状细胞交织而成的髓,具有输导作用。在海带生活史中,无性生殖和有性生殖、孢子体和配子体交替出现,因此有明显的世代交替。海带生长在较清凉的沿海海水中,可供食用、药用和制碘工业原料。

海带
裙带菜
裙带菜藻体呈褐色,叶状,革质, 高50-80厘米,可达1.7米,宽20~40厘米。中肋隆起,两侧羽状裂片;柄部扁圆形,固着器假根状。成熟藻体柄两侧生有木耳状重迭皱褶的孢子叶。生长在风浪较大的低潮线及以下1-2米的岩石上,或低潮带石沼中。生长盛期4~6月,可供食用、药用和制胶工业原料,是重要的养殖种类之一。

裙带菜
墨角菜目代表藻种
鹿角菜目(Fucales)的海藻为薄壁组织,顶端生长;单倍体世代退化为卵子与精子,生活史中的其他时期为二倍体。该目的代表藻种是鹿角菜(Slivetia siliquosa),体大而分枝繁多;在风浪大处,藻体则小而分枝简单。鹿角菜是生长于青藏高原的特有野生藻类,富含营养元素,可改善人体消化功能,对肠胃疾病、糖尿病具积极的食疗作用。
红藻门
红藻门(Rhodophyta)植物体外形多样,除少数是单细胞或群体外,绝大多数为多细胞体,其中有简单的单列细胞或多列细胞组成的丝状体,或由许多藻丝组成的圆柱状、亚园柱状、叶状、囊状或壳状,分枝或不分枝的宏观藻体,其中少数种类钙化。藻体直立或匍匐,基部由假根状分枝丝体或多细胞盘状固着器固着于基质上。由藻丝组成的藻体有单轴型和多轴型两种类型。红藻的生殖类型分为营养繁殖、无性生殖和有性生殖。红藻不同于其他藻类(除蓝藻类外),缺乏具鞭毛的生殖细胞。红藻的绝大多数种类都有三个世代的藻体进行世代交替,即孢子体世代、配子体世代和果孢子体世代。世界广泛分布,包括极地,是水生环境的重要成员。红藻绝大多数为海产,少数生于淡水。海产种类生长的深度可达200米,在潮间带则多生于岩石的背阴处、石缝或石沼中,也有少数喜生于暴露的风浪大的岩石上。淡水产种类大多固着于泉水溪流中岩石上或其他基质上。基于纹孔结构进化和分子系统学观点,将红藻门划分为3个亚门:真红藻亚门(Eurhodophytina)、原红藻亚门 (Proteorhodophytina)和小青藻亚门 (Cyanidiophytina),包含7纲,7000多种。其中红毛菜纲和真红藻纲隶属真红藻亚门。
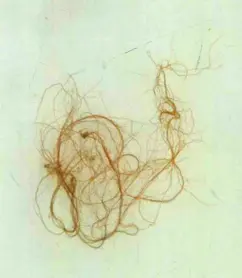
红毛菜
红毛菜目代表藻种
红毛菜目(Bangiales)的藻体在微观的丝状体阶段具有纹孔连接,在宏观阶段则没有纹孔连接。该目紫菜属(Porphyra)中的各藻种如边紫菜(Porphyra marginata)、刺边紫菜(Porphyra dentimarginata)、深裂紫菜(Porphyra schistothallus)等,是广布于世界各地的冷水性潮间带海藻。均具有叶状体从固着器上长出的特征,并且叶状体由1或2层细胞组成。另外,紫菜可用于鲜食或制成干品,干紫菜是市场上畅销的高级副食品。
红盾藻目代表藻种
红盾藻目(Erythropeltidales)的藻体附生在大型藻体上,多细胞,细胞呈盘状、丝状或叶状,细胞含星形或侧壁状色素体,基部细胞有分化,但不产生假根。丝状的或叶状的藻体是居间生长,盘状的藻体是顶端生长。该目红枝藻属(Erythrocladia Rosenvinge)在中国海域只报道有不规则红枝藻(Erythrocladia iregularis)1个物种。不规则红枝藻的藻体附生在大型海藻藻体上,丝状,紫红色。开始藻丝是互相分开的,原始的藻丝向相对的方向生长,这些分枝不断地再进行分枝,表面可见全方位的不规则分枝。
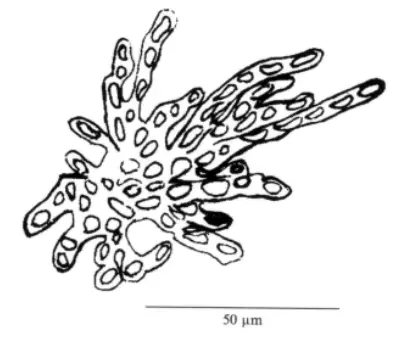
不规则红枝藻
蓝藻门
蓝藻门又称蓝菌门、蓝藻门。一群形态各异的光合自养型细菌,代表看细菌主要系统发育门中的一支,与革兰氏阳性菌亲缘关系较远。是地球上第一种释放氧气的光合微生物,使大气从无氧转变成有氧。叶绿素a 与蓝细菌产生一种称作藻蓝素的藻胆素,使蓝细菌呈蓝绿色,也有的类群产生一种藻红素的藻胆素,通常是红色的或棕色的。蓝细菌广泛分布于自然界的陆地、淡水及海洋中。它们比其他藻类更能耐受不良环境,通常在热泉、盐湖及其他恶劣环境中,蓝细菌是其主要的光养生物。在营养丰富的淡水湖,蓝细菌会大量繁殖形成水华。
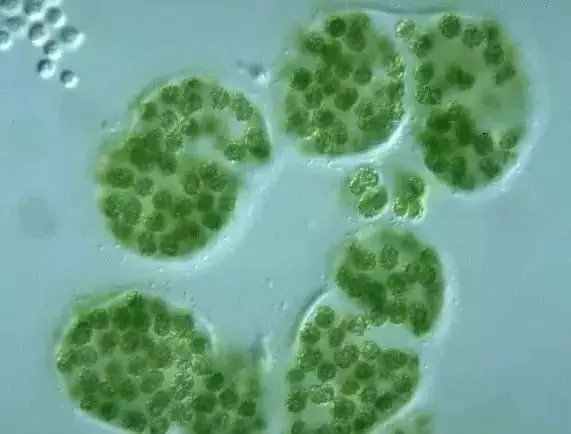
蓝藻门的藻类
念珠藻目代表藻种
念珠藻目(Nostocales)是具有异形胞的丝状体蓝藻。该目藻种的繁殖方式通常通过段殖体;异形胞和(或)厚壁孢子在基部,一种无色发丝在丝状体的顶端;发丝是表皮毛状体的区域,该区域细胞狭长,高度空泡化,几乎呈无色。该目的代表藻种如普通念珠藻(Nostoc commune)球状念珠藻(Nostoc sphaeroides)、圆球念珠藻(Nostoc sphaericum)、浮游念珠藻(Nostoc planktonicum)等均归属于念珠藻属(Nostoc)。另外,该目普通念珠藻,俗称地木耳,是生态系统中的主要先驱物种,可在大部分严苛的环境中生长;在天然药物和制剂制作中有较高的应用价值。
念珠藻目的藻类
色珠藻目代表藻种
色球藻目(Chroococcales)通常形成群体,单细胞的种类少。群体呈球形、椭圆形、不规则形、平板状、穿孔状等。单细胞多呈球形、椭圆形,很少长形的,无顶端和基部的分化。该目中铜锈微囊藻(Microcystis aeruginosa Kiietzing)属于微囊藻属(Microcystis),微囊藻属中的蓝藻有不少种类会形成水华。膜状色球藻(C.membraninus(Meneghini)Nageli)、小型色球藻(C.minor(Kutzing)Nageli)、膨胀色球藻(C.turgidus(Kutzing)Nageli)、易变色球藻(C.uarius
A.Braun)和离散色球藻(C.dispersus(Keissler)Lemmermann)属于色球藻属(Chroococcus Nageli)。
其他藻门
绿藻门
绿藻门植物体由真核细胞组成;藻体形态多样,常为单细胞、群体、丝状体、叶状体,游动或静生,细胞内含叶绿体的色素体,细胞主要进行有丝分裂。繁殖方式亦多种多样,无性繁殖和有性繁殖方式较为普遍。生活史有三种类型即单倍体的藻体型、多倍体的藻体型双单倍体的或称单双倍体的藻体型。绿藻门的分类系统尚无定论,根据F.乐利特 (2012)报道,绿藻门至少包含4纲,20旦。核心绿藻类群(Core Chlorophytes)主要包含3纲:绿藻纲(Chlorophyceae)、共球藻纲(Trebouxiophyceae)、石莼纲 (Ulvophyceae)。除此以外,还有较原始的葱绿藻纲 (Prasinophyceae)和较高等的轮藻纲(Charophyceae)。绿藻门从两极到赤道、从高山到平地均有分布。绝大多数种类产于淡水,少数产于海水,浮游和固着的均有:此外还有气生的种类,少数种寄生或与真菌共生形成地衣。绿藻门(Chlorophyta)的淡水种类占90%,海水种类仅占10%。包含了蕨藻目(Caulerpales)、绒枝藻目(Dasycladales)、管枝藻目(Siphonocladales)等主要的海水种类,丝藻目(Ulotrichales)和鞘毛藻目(Coleochaetales)主要的淡水种类,以及鞘藻目(Oedogoniales)和双星藻目(Zygnematales)全部的淡水种类。
甲藻门
甲藻门(Pyrrophyta)主要是单细胞生物,除少数种类无细胞壁外,都有厚的细胞壁,称为壳。典型的游动甲藻类由横沟把细胞分成上、下两部分,分别称为上、下椎体,具有两根不同的鞭毛。典型的运动甲藻通常被一条横沟(cingulum)分隔为上壳 (epitheca)和下壳 (hypotheca)。一条纵沟(longitudinal sulcus)垂直于横沟。甲藻最普遍的繁殖方式为细胞分裂,有的种类也可以产生动孢子、似亲孢子和不动孢子,少数种类可以进行有性生殖,为同配式。甲藻门仅有1纲,即甲藻纲 (Dinophyceae),包含了原甲藻目(Prorocentrales)、鳍藻目(Dinophysiales)、多甲藻目(Peridiniales)和裸甲藻目(Gymnodiniales)等20目,约3000种。甲藻门在全球广泛分布,是淡水和海洋浮游生物的重要成员。
甲藻门的藻类
金藻门
金藻门(Chrysophyta)植物体类型多样,单细胞或分枝丝状体,细胞形状有梨形、卵形、球形或椭圆形,能运动或不能运动。运动细胞多具2根不等长或等长的鞭毛,也有具1或3根的,少数类群无鞭毛,但能伸出伪足作变形虫样运动。其细胞裸出,或具以果胶质为基质的硅质鳞片,或具囊壳。能运动的单细胞种类的繁殖方法为细胞纵分裂,除细胞纵分裂外,也常断裂成2个或更多的新群体。此门藻类有独特的无性生殖方式,即原生质形成的静孢子,也叫孢囊,孢囊萌发时,其原生质形成一个新个体。根据金藻门在生长时期体制的类型,其分类系统可分为7纲:金藻纲(Chrysophyceae)、黄群藻纲(Synurophyceae)、硅鞭藻纲(Dictyochophyceae)、海生藻纲(Pelagophyceae)褐枝藻纲(Phaeothamniophyceae)、囊壳藻纲(Bicosoecophyceae)和土栖藻纲(Prymnesiophyceae)。金藻门约有1000种,中国约有200种。金藻门分布较广,多为贫营养型淡水水体种类,也有半咸水和海水种类,一般在较冷的季节生长旺盛。
硅藻门
硅藻门(Bacillariophyta)藻体为单细胞或由细胞彼此连接成链状、带状、丛状,辐射状等群体,浮游或着生。细胞壁多数高度硅质化,形成上、下两壳,以壳环套合形成一个硅藻细胞(称为壳体)。金藻门生殖方式以细胞分裂为主,也有复大孢子、小孢子和休眠孢子及由产生具鞭毛的配子或其他有性繁殖方式。硅藻种类繁多,在陆地、淡水、海水中均有分布。有学者于2004年提出将硅藻门分成2亚门3纲,即圆筛藻亚门(Coscinodiscophytina)的圆筛藻纲(Coscinodiscophyceae),硅藻亚门(Bacillariophytina)的中型硅藻纲(Mediophyceae)和硅藻纲(Bacillariophyceae)。中国普遍沿用金德祥(1978)的分类系统,将硅藻门分为中心纲(Centricae)和羽纹纲(Pennatae)。本门约有300属,约12000种。硅藻是某些浮游动物、贝类、鱼类、鲸类以及其他水生动物的重要饵料。浮游硅藻是海洋中主要的初级生产力。化石硅藻在石油勘探、地层划分和对比以及对古地理、古气候及古生态的研究方面有着重要的科学意义。
硅藻门(Bacillariophyta)包含了圆筛藻目(Coscinodiscales)、根管藻目(Rhizosolenialed)、盒形藻目(Biddulphiales)等目。硅藻种类繁多,其细胞单生或连成带状、链状、辐射状等群体;细胞壁几乎高度硅质化,形成上、下两壳,以壳环带套合形成一个硅藻细胞,亦称为壳体。此外,以硅藻壳体为主形成的硅藻土矿具有重要的经济价值。

硅藻细胞
分布范围
编辑目前已知的藻类种类已超过3万种。藻类广泛存在于地球各类环境中,例如生长于高山雪地的藻类、与裸露岩石地衣共生的藻类,再到生活在荒漠沙土、温泉中的藻类。但藻类主要生活在水中,可以是淡水、半盐水或海水中,如蓝藻、硅藻和绿藻等附植藻类。
生长发育
编辑生长规律
藻类的生命周期较短,生长规律依次可分为停滞期、指数生长期、线性生长期、生长衰减期、稳定期和衰亡期六个阶段,其生长发育规律通常呈“S”型曲线。在停滞期,藻类为适应新环境需进行生理调节,其生长有所延迟;随后藻类在指数生长期不断分裂繁殖,细胞生长速度不断加快,当藻细胞增加到一定数量时,其繁殖速率开始受到光照的限制,呈现出恒定速率的线性生长阶段;随着藻细胞数量的不断增加,营养物质的不足会使其分裂速率逐渐下降至零,此时藻细胞数量不再增加,从衰减阶段进入到稳定阶段;在衰亡阶段,藻细胞会因物质消耗、外部干扰等原因,数量迅速下降。
生活史
具有有性生殖能力的藻类的生活史较为复杂。依据生活史中有几种类型的藻体、体细胞为单倍或二倍体,以及有无世代交替,可分为单体型生活史和双单体型生活史。
单体型生活史中只出现一种类型的藻体,无世代交替现象。根据藻体体细胞的倍性又可分为单体型单倍体生活史和单体型二倍体生活史。如衣藻(Chlamydomonassp)的生活史,衣藻(Chlamydomonassp)细胞是单倍体(n),行有性生殖时藻体细胞直接转化成生殖细胞,仅在合子期为二倍体(2n),合子萌发前经减数分裂,萌发产生新的单倍体藻体(n)。生活史中只有核相交替而无世代交替。
双单体型生活史是指生活史中包含两种个体形态的藻体,即存在世代交替现象。根据两种个体的形态、生活期长短及能否独立生活,又可分为等世代型,不等世代型。红藻门 (Rhodophyta)红毛菜目-(Bangiales) 物种具有典型的异型世代交替生活史,由微型孢子体(2n) 阶段和大型配子体 (n)阶段组成。孢子体经丝状藻丝、壳孢子囊枝、壳孢子形成和放散等阶段产生壳孢子 (2n),壳孢子萌发时进行减数分裂,两次细胞分裂产物形成4个线性排列的单倍体细胞(线性四分子),再经有丝分裂形成配子体。研究证明,壳孢子减数分裂时进行同源重组,但由于线性四分子不分离,因此由基因型杂合的孢子体形成的配子体通常是基因型嵌合的单倍体。
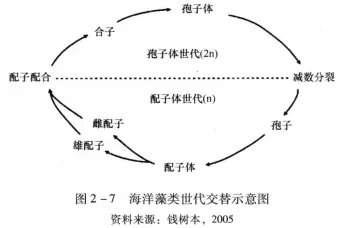
特征特性
编辑形态及细胞结构
藻类的形态和大小差异较大,从有3-10微米大小的藻细胞到70米的巨型水藻。根据有无膜包被的细胞器,藻类的细胞结构可分为原核和真核两种,除蓝藻门和原绿藻门外,其余的藻类均是含有细胞器的真核细胞。原核细胞的主要结构有细胞壁、 细胞质、核糖体,以及由一条裸露的DNA双链构成的拟核,缺少膜包裹的细胞真核细胞通常含细胞壁、质体、线粒体、细胞核、高尔基体和鞭毛等细胞器。
生理特征
藻类的营养方式包括三种:自养、异养和混养。自养藻类通过光合作用将太阳能转变为化细胞物质,以获取获取生长过程中所需的能量;异养藻类的生长模式可分为光能异养、化能异养和光激活异养三种,其中华能异样藻类通常利用有机物作为自身生长的碳源;混养藻类则是指结合自养和异养的生长过程的藻类。
生态习性
藻类作为食物链中的初级生产者,将水、二氧化碳和光转化成有机物,不仅作为食物链的基本食源,而且能形成消费者代谢所必需的氧气。不同藻类对光照、温度的要求不同。自养藻类如绿藻、蓝藻通常需一定的强光照射,其中绿藻对红光和蓝光波长较敏感;而藻类生长温度一般在5-40℃之间。另外,藻类的生长依赖于环境中的碳元素、氮元素和磷元素。藻类对养分的需求量会影响其细胞的生物质组成和生长繁殖过程,藻类可利用二氧化碳或有机碳作为碳源,而氮源和磷源的供给是藻类繁殖的必备条件。
共生特性
藻类和细菌之间存在着普遍的互惠、共生和寄生关系,从单细胞微藻到大型藻类。藻类为共生细菌提供了固定的有机碳,而细菌反过来为藻类提供无机营养物质。例如:藻类和细菌相互作用从而影响地衣的生产;珊瑚可促进藻类进行碳的固定,而藻类能提高珊瑚对环境变化的抗逆性;根瘤菌和固氮菌在促进藻类生长中的重要作用;寄生细菌常存在于藻类细胞壁中并不断促进其降解。

地衣
突变
形态突变
形态突变在藻类中很常见,但大多数藻体形态变异不是真正的突变。因为藻类的形态和颜色很容易受到环境条件的影响而发生改变。即使是同一个物种,如果生活环境不同,它们的形态也会发生较大的变化。但是,有的藻类形态变异体是属于真正的突变体,如藻体变粗、 分枝变短变稀的江蓠,形态变细长而藻体变薄的紫菜,叶片基部变宽大的海带等,它们的形态特征在其有性生殖的后代中仍然能稳定地重现。利用这样的藻类形态突变体,可以培育出优良的新品种。
颜色突变
色素突变体一般在含有多种不同类型色素的藻类中容易出现。例如,坛紫菜含有叶绿素a、藻红色素、藻蓝色素和叶黄素,其色素突变体颜色可以由原来的棕红绿色(野生色) 变成浅红色、浅黄绿色、紫红色等突变颜色。藻类色素突变体可以大致分成自然色素突变体和人工色素突变体,前者是由于自然突变造成的,出现的概率非常低,而后者是由于人工诱变后造成的,出现概率较高。藻类色素突变体不仅颜色和色素含量发生了改变,而且藻体的光合作用效率、生长速度、成熟期、孢子放散量等均会发生改变。
繁殖方式
编辑生殖方式
藻类的繁殖方式可分为无性繁殖和有性繁殖。例如,蓝藻门的颤藻(Oscillatoria)以克隆的破裂方式或段殖体、小繁殖细胞的形成进行无性繁殖;异鞭藻门黄藻纲(Xanthopyceae)的大多数藻类则通过断裂、游动孢子和不动孢子等方式进行无性繁殖,少数黄藻纲藻类如气球藻属(Bortrydium)、黄丝藻类属。(Tribonema)和无隔藻属(Vaucheria)的藻类可进行有性生殖;红藻门海萝属(Gloiopeltis)可通过四分孢子发育成雌雄配子体进行有性繁殖。
无性生殖
无性生殖包括一些单细胞的藻类通过细胞分裂、产生游动孢子、不动孢子或似亲孢子增加个体的数量。一些定型群体,如团藻可通过形成似亲群体增加其群体数量。一些不定群体、丝状体或其他结构较为复杂的多细胞体,如绿藻门的浒苔、红漢门的多管藻等甚至可以通过藻体的断裂增加藻体的个体数量。一些丝状的蓝藻,如颤藻,通过形成藻殖段和藻体的断裂进行生殖。
一些藻类藻体的部分细胞不产生生殖细胞,却可以以多种形式离开母体后继续生长,并能直接发育成新个体,这种繁殖方式为营养繁殖,是无性繁殖的一种方式。新发育成的个体与原母体在外部形态、生长势和细胞结构等方面完全相同。营养繁殖可能发生在配子体世代,也可能发生在孢子体世代,还可能两个世代都存在。在许多情况下,一个配子体或孢子体体细胞通过无性繁殖的细胞群在遗传上是同质的,或者说是一个克隆。营养繁殖可用于扩增藻类生物量,且因藻类可保持很高的营养繁殖速度。
一些藻类雌配子不经过雄配子受精,直接发育成新个体,这种单型生殖的现象称为孤雌生殖。绿藻和褐藻存在孤雌生殖方式已被证实。20世纪70年代初,中国藻类遗传学家方宗熙以海带为研究对象,发现了全新的“雌配子体-孢子体一雌配子体”的生活史循环。
有性生殖
有性生殖中根据相结合的配子形态、大小和行为分为同配、异配和卵配三种类型。有些藻类相结合的两个配子的形态、大小和行为完全一致,如衣藻,为同配生殖。若相结合的两个配子的形态相似,但大小和行为上有差异,如松藻,则为异配生殖;若相结合的两个配子在形态,大小和行为上均表现出明显的差异,如海带,则为卵配生殖。
紫菜属所有种类都进行有性生殖,其雌雄生殖组织分别称为果孢子囊和精子囊群,两者在雌雄同体藻体上的分布分三种情况,第一种为藻体一半是雌性,一半是雄性;第二种为精子囊呈条带状嵌于果孢子囊群中;第三种为果孢子囊与精子囊群不规则混生。研究表明,在实验条件下雌雄同体条斑装菜能够自体受精,其自交率可高达45.8%-55.6%。红毛菜目紫菜属条斑紫菜和圆紫菜(P. suborbiculatakjellm)即为雌雄同体,半叶紫菜(P katadai) 是雌雄同体为主兼有雄性株,而坛紫菜则是雌雄异体为主兼有雄雄同体。
栽培养殖
编辑养分来源
藻类需要提供氮、磷、钾的混合物肥料才能良好生长。有研究发现每吨藻类生物质总共需要40-90公斤氮和3-15公斤磷,而硝酸盐和磷酸盐是藻类生长介质中使用的氮和磷的常见来源。另外,在藻类生长过程中钙、铁和镁等微量元素亦是必需的,主要来源包括农业、工业、动物废水和海水。
培养系统
藻类生长系统主要有露天池塘、光生物反应器和异养生物反应器三种。前两者均利用光合作用的太阳能作为主要的能量来源,而后者则利用还原碳作为主要能量来源。此外,在培养过程中,需维持基本的营养供应、温度的恒定控制;同时,对培养用水多使用海水或但海水混合培养,以减少对淡水资源的依赖。
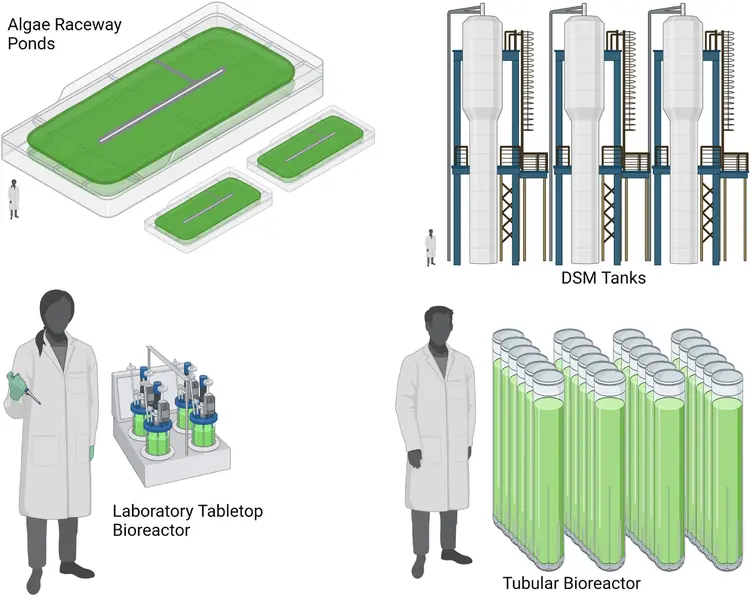
培养系统
具体藻类栽培方法
支柱式栽培
这种栽培方式是一种潮间带主要的紫菜栽培方式。在适当的潮间带滩涂上安设成排的木桩或竹竿作为支桩,将长方形的网帘按水平方向张挂到支柱上。最初编网帘的材料主要是棕绳和细竹条,现在大部分采用维尼纶等化学纤维编织网帘。

支柱式栽培
半浮动式栽培法
它的筏架结构兼有支柱式和全浮动筏式的特点,即整个筏架在涨潮时可以像全浮动式那样经常漂浮在水面上,当潮水退落到筏架露出水面时,它又可以借助短支腿像支柱式那样平稳地架立在海滩上。由于网帘在低潮时能够干露,因而硅藻等杂藻类不易生长,有利于紫菜的早期出苗,而且生长期较长,紫菜质量好。由于网帘经常漂浮水面故能够接受更多的光照,紫菜生长也比较快。因此,半浮动式栽培法已经得到了广泛的应用。
全浮动式栽培
这种栽培方式适合不干露的浅海区栽培紫菜。尤其在冬季有短期封冻的北方海区,还可以将全浮动架沉降到水田以下来度过冰冻期。全浮动栽培的主要缺点是网帘不能及时干露,不利于紫菜叶状体的健康生长和抑制杂藻的繁生。如果网帘的干露问题没有得到解决,用全浮动筏式栽培进行紫菜育苗,常常得不到好的出苗效果。全浮动式栽培还存在着菜体容易老化、叶体上容易附生硅藻、产品质量较差、栽培期较短等问题,单产量也不如半浮动式栽培稳定。
功用价值
编辑藻类在不仅经济方面提供了高生产力、高商品化的应用潜能,还具有作为动物饲料和食物配料的悠久历史,同时在中医药领域也有广泛应用,不仅如此,藻类在生物塑料、生物燃料和污水净化方面的生态价值也十分可观。
经济价值
藻类具有高生物质生产力、高营养价值以及商品规模生产的潜力,已广泛应用于农业生产、食品加工和化妆品工业上。根据世界银行的估计, 500万平方公里的海藻生产面积,约仅占海洋表面的0.3%,但足以产生与全球农业每年生产的生物量一样多的生物量;螺旋藻每公顷产生的生物量是高产杂交玉米的10倍左右。在食品加工方面,大多数以微藻为基础的商业产品的生产商位于亚洲或澳大利亚,且逐年呈现出可观的增长趋势,具较大的市场潜力。另外,大量的商业实体已倾向于在其化妆品中大量使用藻类代谢物,常用作抗氧化剂、防晒霜、保湿剂等,以增强皮肤抗磨损、防晒等能力,例如用于头发治疗的藻类蛋白质、化妆品中的虾青素、瑞士雪藻混合海杏仁油用于抗衰老面霜等等。
食用价值
绝大多数的藻类物种暂未有明确的食用价值研究和评估,而微藻和海藻则已有作为动物饲料和食物配料的悠久历史。微藻和海藻含有丰富的营养成分,如高水平的必需氨基酸、抗氧化物质、维生素和碳水化合物等。利用微藻生物质制成的饲料往往能增强免疫反应、提高疾病耐受性及抗菌和抗病毒作用。而人类往往直接提取微藻衍生的色素和维生素或制成膳食补充剂,以干粉、胶囊等形式直接食用,另外,微藻亦可制成调味剂、增稠剂和甜味剂等配料添加至零食、传统面食和饮料中。海藻的主要市场则是农业和藻胶体行业,如海藻酸和海带等单一产品,可直接出售食用,食品包括寿司用紫菜片、其他干海苔和裙带菜;同时,海藻在食品生产中还可作为可食用包装生物材料。

裙带菜
药用价值
藻类作为中药药材,可以提高和改善人的健康状况。例如,金鱼藻具有清热利水、凉血止血的作用;珊瑚藻、软骨藻和蜈蚣藻均可驱虫,可治疗蛔虫病;螺旋藻有助于益气养血、健脾补肾,常作为病后体虚、贫血、肿瘤化疗及放疗引致免疫功能低下的辅助治疗药。此外,藻类也常作为药品的主要成分,如螺旋藻胶囊、清瘿五海丸等等。
生态价值
生物塑料
藻类衍生物是生物塑料生产的潜在可再生生物质来源,如微藻淀粉、蓝藻衍生的聚羟基烷酸酯、提取自海藻琼脂和藻酸盐等物质,其中藻类中的微藻生物质的共混材料,可提高其机械性能,从而降低断裂伸长率,延长使用寿命,以有效解决传统塑料的不当使用给可持续能源和环境保护所带来问题。另外,藻类尤其是微藻,可通过自身合成的毒素系统或相关酶降解传统塑料材料。
生物燃料
藻类生物质是生产生物燃料的合适原料,是化石燃料的潜在替代品。藻类生物质由藻类细胞通过光照和二氧化碳产生的碳水化合物、蛋白质和脂质组成,属于非消耗性原料,具有较高的生物质生产力,亦具备作为光合作用电子供体和生产各生物产品的潜力。
污水净化
藻类可有效地应用于污染水重金属的生物吸附,且吸附过程中不依赖于温度,在pH值4-7.5的中到弱酸性环境中更有效,以有助于防止二次污染。藻类亦可从城市废水中回收溶解的氮元素和磷元素,而其藻类生物炭可有效地提高土壤肥力,减少农业对有限磷矿物来源的依赖,另外藻类对废水中的微生物亦具有明显的降解作用。

微藻
危害
编辑水体中氮、磷元素等营养盐的增加为藻类提供了有利的繁殖条件,有害藻类在淡水和海水中的爆发性增长现象——水华和赤潮,其引起的问题已是全球关注的问题之一。虽然藻类的尺寸和比重都较小,但其带负电荷,对处理过程中的过滤、混凝效果会有不利影响;另一方面,在处理污染水体的消毒工序中,藻类会与氯作用生成复杂的氯化消毒副产物,降低了饮用水的安全性;此外,传统的水处理方法不能有效去除藻毒素,当长期饮用受到藻类毒素污染的水,可能会引起消化道、肠道等疾病,某些藻类毒素还具有致畸、致癌、致突变作用,若直接接触含藻毒素的水也会造成皮炎和结膜炎等。同时,其爆发时往往会漂浮或悬浮于水面,使水体浑浊,水中溶解氧含量急剧下降,进一步导致水生生物的大量死亡,从而严重影响生态平衡,致使水生生态系统退化、水产养殖破坏和经济损失。
为了减缓有害藻华的发生,可使用生物修复方式如噬菌体、细菌、真菌和浮游动物等,亦有许多学者研究结果表明了化感物质作为植物分泌的次生代谢产物,对藻类具有明显的抑制作用。

赤潮
相关研究
编辑生产技术
藻类具有较高的经济价值、食用价值和生态价值,但技术和经济瓶颈阻碍了藻类的商业用途发展,因此许多学者聚焦于生产技术的优化研究,以期提高工艺效率和降低成本,实现藻类的商业应用。许多学者已着重研究了藻类脂质的提取技术,同时不断优化其生产生物氢、生物甲烷和生物炭等生物基产品的工艺耦合方法,以提高整体的经济性和环保性。另外,许多学者聚焦于藻类生物聚合物的合成方法和生产工艺的研究,而合成方法和生产工艺研究的不断完善,使得藻类生物聚合物在材料工程应用领域上也有望得到拓宽。
纳米技术
藻类介导的纳米颗粒(NPs)在生物医学有较大的应用潜力。许多研究已表明,藻类介导的纳米颗粒(NPs)具一定的抗菌和抗真菌活性,且能有效地抑制细菌生物膜形成从而降低其粘附性,有望作为新型、强效的抗菌剂和抗真菌剂在未来广泛使用;另外,藻类介导的纳米颗粒(NPs)具较好的抗癌活性和生物传感作用,在癌症治疗和靶向递送抗癌药物、检测人体激素的类型和水平特别是癌症诊断方面具较大的应用潜力。
除藻剂影响
除藻剂使藻类群落的均匀度和丰富度得到提升,稳定性得到加强,不容易造成某优势藻种相对丰度过高而再次引发藻华。一项研究表明,生物除藻剂既能控制藻华,又能提高存活藻类群落的稳定性,降低了藻华二次爆发的风险,是一种可改善水生态环境质量的绿色除藻技术。基于PMA前处理的高通量测序技术为研究除藻剂处理后藻华水体中活藻群落结构的响应提供了有效手段。
参考资料
编辑展开[1]R. E. Lee. 藻类学. 2012: 1.
[2]肖培根编;刘德培,王辰总主编. 中华医学百科全书. 2022: 86.
[3]张彦文,周浓主编;马三梅,刘虹,刘建福,吕艳杰,李海生,李景蕻副主编;王鹏,王奇志,汤晓辛等编.. 植物学. 2014: 95-96. (3)
[4]Ścieszka S, Klewicka E.. Algae in food: A general review. Critical reviews in food science and nutrition, 2019[2023-10-15].
[5]水中有机物和营养盐对藻类繁殖的耦合作用研究.中国知网. [2023-10-15].
[6]藻类.中国大百科全书. [2023-10-15].
[7]刘涛. 藻类系统学. 2017: 31. (9)
[8]海萝属藻类繁殖生物学及增养殖研究进展.中国知网. [2023-10-15].
[9]Singh S P, Singh P.. Effect of temperature and light on the growth of algae species: A review. Renewable and sustainable energy reviews, 2015[2023-10-15].
[10]Ramanan R, Kim B H, Cho D H, et al. . Algae–bacteria interactions: evolution, ecology and emerging applications. Biotechnology advances, 2016[2023-10-15].
[11]Diaz C J, Douglas K J, Kang K, et al.. Developing algae as a sustainable food source. Frontiers in Nutrition, 2023[2023-10-15].
[12]Packer M A, Harris G C, Adams S L.. Food and feed applications of algae. Algae Biotechnology: Products and Processes, 2016[2023-10-15].
[13]国家中医药管理局《中华本草》编委会. 中华本草. 1999: 1. [2023-10-15].
[14]药智数据中成药处方.药智数据. [2023-10-15].
[15]Dang B T, Bui X T, Tran D P H, et al.. Current application of algae derivatives for bioplastic production: A review. Bioresource Technology, 2022[2023-10-15].
[16]Chia W Y, Tang D Y Y, Khoo K S, et al.. Nature’s fight against plastic pollution: Algae for plastic biodegradation and bioplastics production. Environmental Science and Ecotechnology, 2020[2023-10-15].
[17]Yaashikaa P R, Devi M K, Kumar P S. . Algal biofuels: Technological perspective on cultivation, fuel extraction and engineering genetic pathway for enhancing productivity. Fuel, 2022[2023-10-15].
[18]Lin Z, Li J, Luan Y, et al.. Application of algae for heavy metal adsorption: A 20-year meta-analysis. Ecotoxicology and Environmental Safety, 2020[2023-10-15].
[19]Cole A J, Paul N A, De Nys R, et al. . Good for sewage treatment and good for agriculture: Algal based compost and biochar. Journal of environmental management, 2017[2023-10-15].
[20]Mathew M M, Khatana K, Vats V, et al. . Biological approaches integrating algae and bacteria for the degradation of wastewater contaminants—a review. Frontiers in Microbiology, 2022[2023-10-15].
[21]Li B, Yin Y, Kang L, et al. . A review: application of allelochemicals in water ecological restoration——algal inhibition. Chemosphere, 2021[2023-10-15].
[22]单细胞水生藻类增长的非线性动力学问题.中国知网. [2023-10-15].
[23]饮用水源水中藻类污染危害及检测研究现状.中国知网. [2023-10-15].
[24]Balaji-Prasath B, Wang Y, Su Y P, et al.. Methods to control harmful algal blooms: A review. Environmental Chemistry Letters, 2022[2023-10-15].
[25]Pal M, Yesankar P J, Dwivedi A, et al.. Biotic control of harmful algal blooms (HABs): A brief review. Journal of environmental management, 2020[2023-10-15].
[26]Sibbald S J, Archibald J M.. Genomic insights into plastid evolution. Genome biology and evolution, 2020[2023-10-15].
[27]Mendes M C, Navalho S, Ferreira A, et al.. Algae as food in Europe: An overview of species diversity and their application. Foods, 2022[2023-10-15].
[28]Wang X, He L, Ma Y, et al. . Economically important red algae resources along the Chinese coast: History, status, and prospects for their utilization. Algal Research, 2020[2023-10-15].
[29]藻类体细胞育苗.中国大百科全书. [2023-10-15].
[30]王素娟. 海藻生物技术. 1994: 106-107. (3)
[31]藻类耐高温性状选育.中国大百科全书. [2023-10-15].
[32]褐藻门.中国大百科全书. [2023-10-15].
[33]蓝细菌门.中国大百科全书. [2023-10-15].
[34]红藻门.中国大百科全书. [2023-10-15].
[35]中国科普博览——植物博物馆.中国科普博览. [2023-10-15].
[36]调味料及添加剂对裙带菜加工色泽的影响.中国知网. [2023-10-15].
[37]热带西太平洋常见浮游植物.科学文库. [2023-10-15].
[38]鹿角菜多酚提取工艺优化及抗氧化活性研究.中国知网. [2023-10-15].
[39]藻类.中国生物志库. [2023-10-15].
[40]钱树本主编. 海藻学. 2014: 111-112. (5)
[41]普通念珠藻的基因组组装与进化分析.中国知网. [2023-10-15].
[42]王赛,杨扬,胡丹心著. 流溪河流域水环境与浮游生物调查研究. 2020: 61. (2)
[43]绿藻门.中国大百科全书. [2023-10-15].
[44]甲藻门.中国大百科全书. [2023-10-15].
[45]金藻门.中国大百科全书. [2023-10-15].
[46]硅藻门.中国大百科全书. [2023-10-15].
[47]中国附植藻类的地理分布格局、群落结构及与环境的关系.中国知网. [2023-10-15].
[48]赵淑江. 海洋藻类生态学. 2014: 33.
[49]藻类雌雄同体单倍体.中国大百科全书. [2023-10-15].
[50]藻类形态突变体.中国大百科全书. [2023-10-15].
[51]藻类色素突变体.中国大百科全书. [2023-10-15].
[52]藻类生殖生物学.中国大百科全书. [2023-10-15].
[53]蔡生力主编. 水产养殖学概论. 2015: 152-153. (3)
[54]Vigani M, Parisi C, Rodríguez-Cerezo E, et al. . Food and feed products from micro-algae: Market opportunities and challenges for the EU. Trends in Food Science & Technology, 2015[2023-10-15].
[55]Ullmann J, Grimm D. . Algae and their potential for a future bioeconomy, landless food production, and the socio-economic impact of an algae industry. Organic Agriculture, 2021[2023-10-15].
[56]Joshi S, Kumari R, Upasani V N. Applications of algae in cosmetics: An overview. Int. J. Innov. Res. Sci. Eng. Technol, 2018[2023-10-15].
[57]Kusmayadi A, Leong Y K, Yen H W, et al.. Microalgae as sustainable food and feed sources for animals and humans–biotechnological and environmental aspects. Chemosphere, 2021[2023-10-15].
[58]Olaniran A F, Akinsanola B A, Taiwo A E, et al. . Algae Utilization as Food and in Food Production: Ascorbic Acid, Health Food, Food Supplement and Food Surrogate. Next‐Generation Algae: Volume I: Applications in Agriculture, Food and Environment, 2023[2023-10-15].
[59]Mendes M C, Navalho S, Ferreira A, et al. . Algae as food in Europe: An overview of species diversity and their application. Foods, 2022[2023-10-15].
[60]Ahmad A, Banat F, Alsafar H, et al. . Algae biotechnology for industrial wastewater treatment, bioenergy production, and high-value bioproducts. Science of The Total Environment, 2022[2023-10-15].
[61]Kumar M, Sun Y, Rathour R, et al. . Algae as potential feedstock for the production of biofuels and value-added products: Opportunities and challenges. Science of the Total Environment, 2020[2023-10-15].
[62]Kartik A, Akhil D, Lakshmi D, et al. . A critical review on production of biopolymers from algae biomass and their applications. Bioresource Technology, 2021[2023-10-15].
[63]Chaudhary R, Nawaz K, Khan A K, et al.. An overview of the algae-mediated biosynthesis of nanoparticles and their biomedical applications. Biomolecules, 2020[2023-10-15].
[64]Koyande A K, Chew K W, Manickam S, et al.. Emerging algal nanotechnology for high-value compounds: A direction to future food production. Trends in Food Science & Technology, 2021[2023-10-15].
[65]改良高通量测序技术揭示除藻剂对藻类群落的影响.中国知网. [2023-10-15].
内容由百科小编提供,本内容不代表globalbaike.com立场,内容投诉举报请联系全球百科客服。如若转载,请注明出处:http://globalbaike.com/15502/