黑猩猩(学名:Pan troglodytes)是生活在赤道非洲,中等体型,用指关节行走的类人猿。黑猩猩身上被又长又粗的毛发覆盖,颜色为深棕色至黑色,下颌突出,眉脊明显。面部和前额中心主要裸露,头部的其他部分被毛发覆盖。手长而细,前肢略长于后肢,没有尾巴。此外,黑猩猩长着锋利的犬齿。成年黑猩猩平均肩高(站立)为150厘米。成年雄性体长(四肢着地,从鼻子到臀部长度)为77~92.5厘米,雌性为70~85厘米。成年雄性体重为28~56千克,雌性为20~46千克。
分类与演化
编辑国际自然保护联盟(IUCN)将黑猩猩分为四亚种:西非亚种、尼日利亚—喀麦隆亚种、中非亚种、东非亚种。
西非黑猩猩,也称为上几内亚黑猩猩(P. t. verus),分布于塞内加尔东南部和马里南部,东南至达荷美峡谷(贝宁)或尼日尔河。西非黑猩猩的下巴经常长有浓密的白色“胡须”,眼周为深色。成年后,脸上一般带点粉色。脸较宽呈灰白色,并比较突出,前额较窄,耳朵大。头发沿着头部中线分开。手掌和脚底为灰白,手指上有不规则的深色斑点;
尼日利亚—喀麦隆黑猩猩,也称为埃利奥特或几内亚湾黑猩猩(P. t. ellioti),分布于尼日利亚南部和喀麦隆西部,可能从达荷美峡谷(贝宁)或尼日尔河以南到塞内加尔河下游。与西非亚种相比,尼日利亚—喀麦隆黑猩猩耳朵更小,更靠近头部,头顶更圆,眉脊更直,身材更纤细,成年时脸、手和脚均为黑色;
中非黑猩猩,也称为下几内亚黑猩猩(P. t. troglodytes),分布于塞内加尔河东南至乌班吉河,南至刚果河。成年后的皮肤,包括脸、耳朵、手掌和脚底,通常为深棕色或黑色,耳朵小到中等,前额经常形成明显的秃斑;
东非黑猩猩,也称为长毛黑猩猩(P. t. schweinfurthii),分布于乌班吉河,东跨刚果民主共和国,刚果河北,卢阿拉巴河东至坦桑尼亚西南部。体型比其他亚种稍小,毛发长,尤其在脸和肩膀周围。成年后脸上为浅棕粉色到棕色或灰黑色,有些个体的嘴唇附近有明显的粉红色痕迹,脸通常比西非黑猩猩长,手掌和脚掌通常为砖红色至青铜色。
DNA证据表明,不到100万年前,黑猩猩与倭黑猩猩分化。遗传研究学表明,约800~600万年前的晚中新世,黑猩猩与人类类群分化。
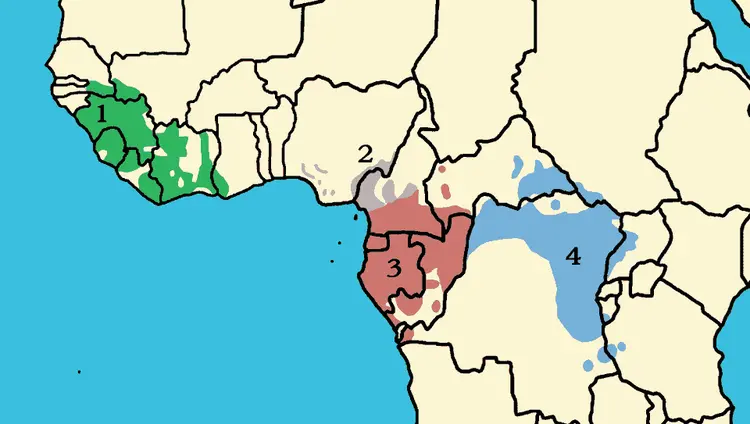

黑猩猩四亚种分布,1为西非黑猩猩,2为尼日利亚—喀麦隆黑猩猩,3为中非黑猩猩,4为东非黑猩猩
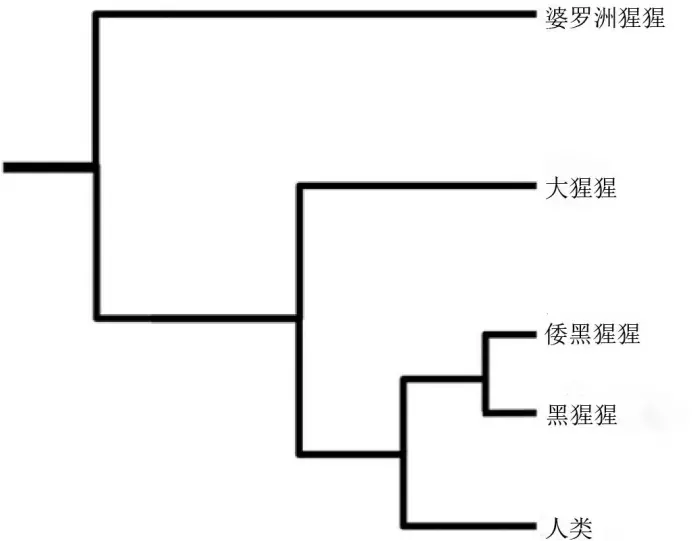
灵长类关系图,显示黑猩猩的位置
形态特征
编辑成年黑猩猩雄性体长(四肢着地,从鼻子到臀部长度)为77~92.5厘米,雌性为70~85厘米。成年黑猩猩平均肩高(站立)为150厘米。成年雄性体重为28~56千克,雌性为20~46千克。

黑猩猩是体型适中,体格健壮,用指关节行走的类人猿。黑猩猩身上被又长又粗的毛发覆盖,颜色为深棕色至黑色。许多老年黑猩猩的下背部、腿部、下巴有浅棕色或灰色毛发。未成年黑猩猩的肛门上方有一簇白毛。黑猩猩两性颜色类似,但成年雌性肩膀较窄,体重约为成年雄性的80%。幼儿黑猩猩的脸、耳朵、手和脚为粉色,成年通常变成棕色或黑色。
黑猩猩下颌突出,眉脊明显。面部和前额中心主要裸露,头部其他部分被毛发覆盖。虹膜呈棕色到深棕色。巩膜为棕色(罕见为白色)。耳朵完全或部分裸露,与人类的耳朵类似,但更大,可以面朝前方或平放在头部两侧。嘴巴灵活结实。手长而细,手指、手掌、脚掌、脚趾都没有毛发。前肢略长于后肢,没有尾巴。此外,黑猩猩的门齿较大,犬齿锋利,雄性犬齿比雌性大25%~40%。

黑猩猩的门齿
分布栖息
编辑地理分布
黑猩猩是迄今为止地理分布最广的类人猿,分布面积超过260万平方千米。它们只在赤道非洲分布,从塞内加尔南部穿过刚果河以北的森林带,到坦桑尼亚西部和乌干达西部,在13°N和7°S之间不连续分布。分布国家包括25个,如安哥拉、布隆迪、喀麦隆、加蓬、卢旺达,可能在贝宁、布基纳法索和多哥灭绝,即将从塞内加尔、加纳灭绝,在大多数国家的大片地区已消失。
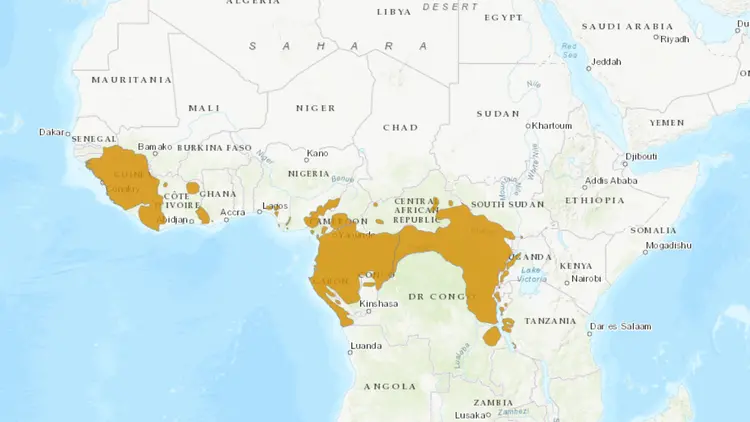
黑猩猩分布
栖息地
黑猩猩大部分栖息地比较潮湿,主要栖息于常绿湿润林或半落叶林、沼泽林、长廊林、林地、再生林以及草地。黑猩猩喜欢成熟的森林栖息地,但也经常在再生林出现。从海平面到最低2949米都可以看到黑猩猩身影,年均降水量一般超过1700毫米。少数种群生活在比较干旱,开阔的草地及林地栖息地。
生活习性
编辑昼夜节律
黑猩猩是昼行性,半陆地动物,每天活跃10~13小时。晚上,黑猩猩主要在树上搭建的巢穴休息睡觉,白天可能也会在巢穴内休息。某些黑猩猩部落可能在地上搭建巢穴。巢穴平均高10~20米,种群内存在很大差异(如,在乌干达布东戈森林,黑猩猩的巢穴达2.5~32.5米高)。雄性的巢穴一般低于雌性,可能因为雄性更重。黑猩猩一般独占一个巢穴,除了带着未独立孩子的雌性。

黑猩猩搭建的树上巢穴
觅食行为
黑猩猩是杂食动物,但主要吃成熟果实,成熟果实约占它们所食食物的60%。黑猩猩每天花55%时间觅食,约14%的时间行走。叶子、果核是主要补充性食物,尤其在成熟果实可利用度较低的月份。黑猩猩也会食用少量未成熟的果实、种子、鲜花、树皮、木头、嫩枝、树液、蜂蜜、昆虫、蛋。
除此之外,黑猩猩还会吃肉,包括中小型有蹄类、鸟类、啮齿类、两栖类等动物。黑猩猩偶尔捕食其他灵长类动物,包括各种猴子、狒狒等,红疣猴是黑猩猩捕食最多的哺乳动物。在极少数情况下,黑猩猩会同类相食。
捕猎几乎由成年雄黑猩猩完成,有时雌黑猩猩与青少年黑猩猩密切跟踪狩猎,并杀死丧失能力的动物。黑猩猩经常与潜在猎物偶遇,但捕猎成功概率与季节性食物的可利用度、成年雄黑猩猩数量,及阻止猎物逃跑,低矮或倒下的树冠相关。捕猎技术高超的雄黑猩猩在场,有利于提高其他雄黑猩猩的捕猎积极性。
黑猩猩一般在黎明不久后开始进食,中午减少,傍晚筑巢前继续。果实倾向于早些时候食用,像叶子这种低质量的食物一般晚些时候食用。
黑猩猩有时与同伴分肉,但很少分给年幼的个体,后者一般从妈妈手中获得。有人认为黑猩猩分肉用于维系社会关系或获取交配机会,但研究表明,分肉是持有者无法容忍同伴密集乞讨、骚扰行为的无奈之举。
黑猩猩会利用工具觅食,曾有研究者观察到它们用树枝钓蚂蚁与白蚁、用木棒挖蜂蜜,用石头和木棍砸坚果、用叶子和苔藓喝水。

东非黑猩猩从狒狒手中抢到一块肉

黑猩猩利用树枝吃蚂蚁
社会行为
黑猩猩是群居动物,黑猩猩群主要由多只成年雄黑猩猩与雌黑猩猩及小黑猩猩组成,黑猩猩群也称为部落。大部分黑猩猩部落包括20~60只黑猩猩,有时超过140只,在受严重干扰的栖息地,有时不到20只。
黑猩猩的家域面积为6~32平方千米,在密度较低的情况下,家域面积超过200平方千米。雄性个体间的家域面积较大,并广泛重叠,雌性经常限制在群家域内狭小的核心区域。在坦桑尼亚贡贝国家公园,黑猩猩的家域面积与雌黑猩猩的繁殖率正相关,表明雄黑猩猩为了养育后代,扩张领域。
雄黑猩猩是高度领域性动物,是黑猩猩社会组织的核心,负责巡逻领域,保护群成员,搜寻食物。雄性间存在社会等级,成年雄性统治所有雌性及小黑猩猩。雄黑猩猩经常组成小群巡逻领域边界地区,一旦发现陌生成员就会尖叫,如果有明显的数量优势,就会袭击,甚至杀死对方。带崽与小型部落的黑猩猩一般避开巡逻黑猩猩群。

乌干达黑猩猩群
生长繁殖
编辑繁殖
黑猩猩发情周期约为35天,发情持续10~12天,雌黑猩猩发情时肛门、阴唇肿胀,怀孕有时也会出现性肿胀。在食物资源丰富时,黑猩猩更容易发情。雄黑猩猩的阴茎呈长形丝状,勃起时长10~18厘米;睾丸较大,平均重158克,能产生大量精子。雄黑猩猩射精后,一部分凝结在“生殖道”中形成精子塞,可以阻止另一只雄黑猩猩的精子进入。
黑猩猩在性肿胀最大时交配,性肿胀与繁殖力、性吸引力相关。雌黑猩猩发情期的最后6天排卵最多,怀孕几率最高,与成年雄性及黑猩猩首领的交配频率增加,有时可能与邻居部落中的雄性交配。
黑猩猩全年繁殖,没有严格的季节性,但长期繁殖模式显示黑猩猩分娩时间的峰值与发情雌黑猩猩数量相关。雌黑猩猩分娩前经常独处,变得无精打采,可能退出社交活动,一般在树上的巢穴分娩。
黑猩猩妊娠期约为230天,一胎一仔,少数双胞胎。野生雌黑猩猩的繁殖年龄超过40岁,最大超过50岁。圈养雌黑猩猩最大繁殖年龄超过60岁。

雌黑猩猩带着孩子
生长发育
黑猩猩出生时体重约1.7千克,之后主要由妈妈负责养育,教导生存技能。黑猩猩刚出生时体弱,1月大时一直紧紧抓住妈妈的肚子,前2个月大时还无法独自站立。5~6个月大时,它们会骑在妈妈的背上。2岁时,小黑猩猩能够独立移动和坐着,并开始移动到妈妈够不到的地方。

黑猩猩妈妈抱着一只八周大的黑猩猩幼崽
黑猩猩发育包括学习使用工具。小黑猩猩在2~4岁时,开始学习使用树枝、木棒钓白蚁,木棒挖蜂蜜,石头砸坚果等,直到几年后熟练掌握。雌性花费很长时间观察妈妈使用工具,并经常练习,雄性则更喜欢与其他黑猩猩进行社交游戏,因此雌性掌握更快。
4~6岁时,黑猩猩断奶,幼儿期结束。6~8岁,黑猩猩进入青少年期,此时仍与妈妈关系亲密,但与群成员的互动增加。8岁后,黑猩猩进入青春期,青春期的黑猩猩在社交互动上有所改变。它们开始离开妈妈,在不同的小群中行走。雌黑猩猩约在8岁时,开始出现微弱、无规律性的性肿胀,10~11岁时性肿胀变得强烈、更有规律性。青春期的雌黑猩猩与无亲缘关系的雄黑猩猩交往更频繁,第一次完全肿胀后迁入新的黑猩猩部落。
大部分雌黑猩猩扩散,离开出生群,少数仍留在出生群。雌黑猩猩一旦迁入新的黑猩猩部落,一般不再更换,但少数仍会更换,约14岁时繁殖第一胎,有些西非黑猩猩可能早至9~10岁。雄黑猩猩的青春期一般始于8岁左右,肌肉、睾丸快速增长,攻击性行为增加,成为成年雄黑猩猩的跟班,开始参与狩猎、巡逻等活动。15岁左右,进入青春期末期,雄黑猩猩统治所有雌黑猩猩及青少年黑猩猩。雄黑猩猩一般留守出生群。
野生黑猩猩一般可活50岁,最大寿命可能超过60岁,某些圈养黑猩猩被认为超过70岁,例如著名的圈养黑猩猩“奶酪”。

坦桑尼亚玛哈尔国家公园的黑猩猩群
种间关系
编辑除了狩猎外,黑猩猩很少与其他动物互动,偶尔与其他灵长类(如狒狒)互相梳毛、玩耍,有时被其他猴子袭击。
许多黑猩猩种群似乎没有受到天敌的干扰,但潜在捕食者包括豹子、狮子、尼罗鳄。豹子袭击是科特迪瓦塔伊森林黑猩猩死亡的主要原因,一项三年研究发现,豹子捕食占黑猩猩死亡数量的39%。在坦桑尼亚玛哈尔(Mahale)国家公园,一个过路狮群至少捕食6只黑猩猩。
物种保护
编辑种群现状
2003年,国际自然保护联盟(IUCN)统计黑猩猩数量总计约为172700~229700只。2016年每个亚种估计为:尼日利亚—喀麦隆黑猩猩数量最少,可能不到6000~9000只,东非黑猩猩为181000~256000只,包括刚果民主共和国北部一个大种群,中非黑猩猩约为140000只,西非黑猩猩约为18000~65000只,整体数量呈下降趋势。
保护级别
2016年国际自然保护联盟(IUCN)将黑猩猩列为濒危(EN)物种,所有亚种都列为濒危(EN)物种,2016年列入《濒危野生动植物种国际贸易公约》附录Ⅰ,禁止贸易。
致危因素
黑猩猩主要面临的威胁有盗猎、栖息地丧失与破坏、疾病。
尽管猎杀、捕捉、消费类人猿是非法行为,但盗猎仍旧是黑猩猩的最大威胁。人们为了获取丛林肉、捕捉小黑猩猩作宠物或直接贩卖,而猎杀黑猩猩。有时为了保护作物,人们可能投毒或报复性杀害黑猩猩。
由于农业扩张、木材开采、采矿,黑猩猩栖息地遭到破坏和退化,威胁长期生存。
导致中非黑猩猩数量下降的第3个主要原因是传染病,尤其是埃博拉病毒(EVD)。因为黑猩猩和人类非常相似,黑猩猩可能患有和人类相似的传染病。呼吸道疾病和炭疽病,是一些习惯于人类存在,与人类离得较近的黑猩猩种群死亡主要原因。

刚果民主共和国珍妮古道尔保护区的一只雄黑猩猩
保护措施
1、严格执行野生动物法,有效管理黑猩猩保护区。2016年,国际自然保护联盟(IUCN)将黑猩猩列入《濒危野生动植物种国际贸易公约》附录Ⅰ,禁止贸易。
2、黑猩猩在许多国家公园内出现,但大部分仍在保护区外分布,加强黑猩猩的栖息地保护。2014年,国际自然保护联盟灵长类专家组( Primate Specialist Group)在黑猩猩的地理分布范围内进行科学合理的土地利用规划,以避免破坏黑猩猩栖息地,建立大规模农业、工业开采,尤其是油棕榈种植园。
3、对黑猩猩数量和种群健康进行长期标准化监测。SMART已在大部分黑猩猩分布范围内使用,用于执法与监测,近十年来,人们一直建议采用标准方法来调查和监测类人猿种群,以便于更准确地监测种群数量变化,现在可以对一系列病原体进行无创诊断,如,检测粪便中的埃博拉病毒。
4、进一步研究减轻埃博拉病毒传播和毒效的方法,包括给黑猩猩接种疫苗,以形成一个阻止埃博拉病毒传播的屏障。
参考资料
编辑展开[1]Butynski, T. M., Kingdon, J. & Kalina, J. (eds). Mammals of Africa. London: Bloomsbury Publishing, 2013: Chapter/species profile: e.g. Williamson, E. A. & Butynski, T. M. 2013. Gorilla gorilla Western Gorilla; pp 410–418.
[2]Humle, T., Maisels, F., Oates, J.F., Plumptre, A. & Williamson, E.A. Pan troglodytes (errata version published in 2018). The IUCN Red List of Threatened Species 2016: e.T15933A129038584. 2016: Accessed on 27 July 2022. [2022-12-22].
[3] Leutenegger, W. & Shell, B. Variability and sexual dimorphism in canine size of Australopithecus and extant hominoids. Journal of Human Evolution, 1987, 16: 359–367.
[4] Hladik, C. M. Chimpanzees of Gabon and chimpanzees of Gombe: some comparative data on the diet. In: Primate Ecology: Studies of Feeding and RangingBehaviour in Lemurs, Monkeys and Apes. ed. T. H. Clutton-Brock. New York: Academic Press, 1977: pp. 481–501.
[5]Struhsaker, T. T. The Red Colobus Monkeys: Variation in Demography, Behavior,
and Ecology of Endangered Species. Oxford: Oxford University Press, 2010: 349 pp.
[6]McGrew, W. C. Tools compared: the material of culture. In: Chimpanzee Cultures (eds R. W. Wrangham, W. C. McGrew, F. B. M. de Waal & P. G. Heltne). Massachusetts: Harvard University Press, Cambridge, 1994: pp. 25–40.
[7]Boesch, C. & Boesch, H. Optimization of nut cracking with natural hammers by wild chimpanzees. Behaviour , 1983, 83: 265–286.
[8]Sugiyama, Y. Drinking tools of wild chimpanzees at Bossou. American Journal of Primatology , 1995, 37: 263–269.
[9]Emery Thompson, M., Jones, J. H., Pusey, A. E., Brewer-Marsden, S., Goodall, J., Marsden, D., Matsuzawa, T., Nishida, T., Reynolds, V., Sugiyama, Y. & Wrangham, R. W. Aging and fertility patterns in wild chimpanzees provide insights into the evolution of menopause. Current Biology, 2007b, 17: 2150–2156.
[10]Fischer, A., Wiebe, V., Pääbo, S., & Przeworski, M. Evidence for a complex demographic history of chimpanzees. Molecular biology and evolution, 2004, 21(5): 799-808.
[11]Won, Y. J., & Hey, J. Divergence population genetics of chimpanzees. Molecular biology and evolution, 2005, 22(2): 297-307.
[12]Wrangham, R. W (ed. T. H. Clutton-Brock). Feeding behaviour of chimpanzees in Gombe National Park, Tanzania. In: Primate Ecology: Studies of Feeding and Ranging Behaviour in
Lemurs, Monkeys and Apes. New York: Academic Press, 1977: pp. 503–537.
[13]Goodall, J. Nest building behavior in free ranging chimpanzee. Annals of the New York Adacemy of Sciences, 1962, 102: 455–467.
[14]Maughan, J. E. & Stanford, C. B. Terrestrial nesting by chimpanzees in Bwindi Impenetrable National Park, Uganda (abstract). . American Journal of Physical Anthropology (Suppl.), 2001, 32: 104.
[15]Nishida, T., Hasegawa, T., Hayaki, H., Takahata, Y. & Uehara, S. Meatsharing as a coalition strategy by an alpha male chimpanzee? In: Topics in Primatology: Human Origins (eds T. Nishida, W. C. McGrew, P. Marler, M. Pickford & F. B. M. de Waal). Tokyo: University of Tokyo Press, 1992: pp. 159–174.
[16]Stanford, C. B., Wallis, J., Mpongo, E. & Goodall, J. Hunting decisions in wild chimpanzees. Behaviour , 1994, 131: 1–18.
[17] Wrangham, R. W. & Peterson, D. Demonic Males: Apes and the Origins of Human Violence. Boston: Houghton-Mifflin, 1996: 350 pp.
[18]Reynolds, V. The Chimpanzees of Budongo Forest. Oxford: Oxford University Press, 2005: 352 pp.
[19]Arcadi, A. C. & Wrangham, R. W. Infanticide in chimpanzees: review of cases and a new within-group observation from the Kanyawara study group in Kibale National Park. Primates , 1999, 40: 337–351.
[20]McGrew, W. C., Baldwin, P. J. & Tutin, C. E. G. Chimpanzees in a hot, dry and open habitat: Mt. Assirik, Senegal, West Africa. Journal of Human Evolution, 1981, 10: 227–244.
[21]Brewer-Marsden, S., Marsden, D. & Emery Thompson, M. Demographic and female life history parameters of free-ranging chimpanzees at the Chimpanzee Rehabilitation Project, River Gambia National Park. International Journal of Primatology , 2006, 27: 391–410.
[22]Boesch, C. & Boesch-Achermann, H. The Chimpanzees of the Taï Forest: Behavioural Ecology and Evolution. Oxford: Oxford University Press, 2000: 316 pp.
[23]Pusey, A. E. Inbreeding avoidance in chimpanzees. Animal Behaviour , 1980, 28: 543–552.
[24]Boesch, C. The effect of leopard predation on grouping patterns in forest chimpanzees. Behaviour, 1991, 117: 220–242.
[25]Tsukahara, T. Lions eat chimpanzees: the first evidence of predation by lions on wild chimpanzees. American Journal of Primatology, 1993, 29: 1–11.
内容由百科小编提供,本内容不代表globalbaike.com立场,内容投诉举报请联系全球百科客服。如若转载,请注明出处:https://globalbaike.com/20315/