硅藻门(Diatomeae,或Bacillariophyta),是一类微小的单细胞真核生物,统称硅藻,在《中国生物物种名录》2023版中,硅藻门归属于色素界。硅藻的物种多样性较高,截止2023年,根据AlageBase网站的统计,已知硅藻的种类名称词条约1.7万种,但其中有很多为基原异名和无效命名,被公认种类仅一万种左右。硅藻分布较广,几乎所有水体,如海洋、河口半咸水、江河等水体中都有硅藻的存在。
历史演化
编辑由于硅藻壁中存有硅质,硅蘖的化石得以保存完好。研究表明最早的硅藻出现在白垩纪,在上白垩纪和第三纪时发展最盛,大量海相硅藻土沉积就来源于白垩纪时期。经历漫长的历史进化后,硅藻仍停留在单细胞及单细胞连接成的群体阶段,硅藻创造了其他藻类中完全找不到的细胞形式,但也因为这一细胞形态,限制了硅藻的继续进化发展。
根据硅藻所含的色素,推测其与金藻、褐藻的亲缘关系较为密切,它们的载色体都含有叶绿素
、叶绿素
和墨角藻黄素。但硅藻特殊的细胞结构及运动方式又与金藻门、褐藻门有很大区别。从细胞壁的构造上,认为硅藻门与黄藻门相近,它们都是由两个半片套合组成的,但两者营养体的倍性又完全不同,硅藻为二倍体,是具叶绿素
、叶绿素
的类群中较特殊的一类。硅藻营养体没有鞭毛,但某些种类生殖细胞具鞭毛,如中心藻纲直链藻属(Melosira)出现有鞭毛的雄配子。又如根管藻属(Rhizosolenia)和菱形藻属(Nitzschia)的细胞分裂时出现伸缩泡、胞口、胞咽等鞭毛藻类所具有的结构,因而设想硅藻可能是从含有叶绿素
、叶绿素
的具鞭毛藻类发展而来的。
分类
编辑分类历史
世界分类史
在18世纪中期,人们开始对硅藻有了初步的认识。贝克(Baker)在其著作《Employment for the Microscope》中描述了两种“动物”,这些被后来认定为硅藻。然而,直到18世纪后半期,人们才开始使用拉丁文对硅藻进行正式命名和描述。1791年,格梅林(Gmelin)创立了Bacillaria(棍形藻属)这一属名,这是第一个正式确立的硅藻属。进入19世纪,硅藻分类研究迎来了快速发展时期。特别是在1825至1896年间,全球范围内共报道了近500个硅藻新种。然而,由于一些硅藻具有单细胞生活且具备运动能力,这导致关于“硅藻是什么”的问题在19世纪仍备受争议。在1844年,库津(Kützing)在其专著指出,无论是否具有运动能力,单细胞或是群体,都应将硅藻定为植物。
1872年,美国硅藻学者汉弥尔顿·史密斯(Hamilton L. Smith)发表了一个分类系统,这一系统被多数学者广泛采纳。史密斯根据无壳缝、具拟壳缝和有壳缝的特征,将硅藻分为三大类和十五小类,为硅藻的分类研究奠定了重要基础。到了1896年,法国硅藻学工作者范·赫尔克(H. van Heurck)进一步将硅藻作为一个科进行分类,他将其分为三个亚科,但大类仍然沿用了汉弥尔顿·史密斯的分类方法。随着研究的深入,卡斯滕(Karsten)在1928年将硅藻作为门,门内仅设一个纲,纲下则分为两个目。1930年,胡斯特德特(Hustedt)将硅藻门的硅藻纲细分为羽纹目(Pennales)和中心目(Centrales)。在羽纹目下,他进一步划分了无壳缝亚目(Arphidineae)、拟壳缝亚目(Raphidioidineae)和单壳缝亚目(Monoraphidineae)等。这一分类系统受到了后人的广泛接受,并在后续的硅藻研究中发挥了重要作用。
随着显微观察技术的不断进步,科尔贝(Kolbe)在1943年首次利用透射电子显微镜(TEM)观察了硅藻的形态特征。1972年,哈斯勒(Hasle)等通过扫描电子显微镜(SEM)发现了硅藻的新结构——唇形突,随后结合新的结构对一些种类进行了分类地位的调整。进入上世纪90年代,朗德(Round)等学者于1990年对硅藻进行了系统的分类,将所有硅藻类群划分为287个属。他们结合SEM和TEM的观察结果,详细论述了分属的依据,这一分类系统至今仍被主流硅藻研究者广泛接受。2004年,梅德林(Medlin)和卡茨马斯卡(Kaczmarska)基于109种硅藻的18S和16S核糖体RNAs基因序列研究,对硅藻的分类系统进行了修订,并对硅系统的演化进行了重建。他们将硅藻门分为两个新的进化分枝,即圆筛藻亚门和硅藻亚门,分别包括1纲圆筛藻纲和2个纲中型硅藻纲和硅藻纲。进入21世纪后,硅藻分类研究继续取得新的进展。克拉默(Krammer)和科西奥莱克(Kociolek)等学者相继报道了许多新的硅藻属和种。尽管高等植物和一些低等植物的分类研究已经进入分子和基因时代,但目前硅藻分类研究的主流仍然是基于形态学特征来进行。
中国分类史
中国硅藻分类的研究起步较晚。俄国硅藻研究者斯克沃尔佐夫(Skvortzov)是第一位报道中国淡水硅藻的学者,根据相关统计斯克沃尔佐夫(Skvortzov)发表了五十多篇与硅藻研究有关的论文。20世纪30年代中期至50年代,以王志稼等人为主要代表,中国的藻类研究逐渐深入,但对于硅藻的专门研究尚未开展。新中国成立之后,湖泊和海洋调查逐步开展,给硅藻的分类研究提供了机会。1961至1976年,中国科学院首次对西藏地区的硅藻进行研究调查,此项研究中发现了44属906个硅藻分类单位,其中新种26个,中国新纪录259个,这也是中国首次比较全面和系统的区系硅藻分类研究,研究结果于2000年发表在《中国西藏硅藻》。中国的硅藻分类研究由金德祥教授于1978年发起,他将硅藻分为2纲9目,即中心纲和羽纹纲,与Hustedt分类系统基本保持一致。20世纪90年代,中科院在全国范围内进行了三次较大范围的藻类生物资源调查,相关研究者先后出版了《中国淡水藻志》系列专著。胡鸿钧等在2006年发表的《中国淡水藻类——系统、分类及生态》也遵循了同样的分类标准。而在《中国生物物种名录》2023版中,硅藻门归属于色素界。
分类争议
硅藻分类系统经历了较长的历程,直至现在也尚存在有争议的不同的分类系统。截止2023年,硅藻通常被视为一个门或者一个纲,而硅藻的拉丁名和下级分类也存在了修订和调整。目前较为常用的分类依据是克拉默(Krammer)和兰格-贝尔塔洛特(Lange-Bertalot)所建立的硅藻区系和朗德(Round)等以扫描电镜研究结果为主要依据的硅藻分类著作。克拉默和兰格-贝尔塔洛特,提出将硅藻分成1纲2目4亚目,即硅藻纲(Class Bacillariophyceae),下设中心目(Order Centrales)和羽纹目(Order Pennales)。Round等将硅藻分为三大纲:圆筛藻纲(Coscinodiscophyceae)、脆杆藻纲(Fragilariophyceae)和硅藻纲(Bacillariophyceae)。
下属分类
国际分类
鉴于截止至2024年,硅藻也尚存在有争议的不同的分类系统,因此此处呈现常用的克拉默(Krammer)和兰格-贝尔塔洛特(Lange-Bertalot)所建立的以欧洲硅藻为主体的区系。
硅藻纲(Bacillariophyceae)
1.中心目(Order Centrales)
圆筛藻亚目(Coscinodiscineae)
根管藻亚目(Rhizosoleniineae)
2.羽纹目(Order Pennales)
无壳缝亚目(Araphidineae)
具壳缝亚目(Raphidineae)
中国分类
根据《中国生物物种名录》2023版数据,硅藻的具体分类如下:
硅藻门(Diatomeae)
1.中心纲(Class Centricae)
盒形硅藻目(Order Biddulphiales)
盘状硅藻目(Order Discoidales)
管状硅藻目(Order Rhizosoleniales)
舟辐硅藻目(Order Rutilariales)
2.羽纹纲(Class Pennatae)
曲壳藻目(Order Achnanthales)
等片藻目(Order Diatomales)
短缝藻目(Order Eunotiales)
舟形藻目(Order Naviculales)
褐指藻目(Order Phaeodactylales)
双菱藻目(Order Surirellales)
形态构造
编辑体型
藻体为单细胞,或以细胞分泌的胶质将细胞连成丝状、带状、星状、放射状等群体。细胞形态多种多样,有圆形、椭圆形、多角形、纺锤形、“
”形、新月形、楔形、棒形、弓形等。
细胞壁
硅藻具有高度硅质化的细胞壁,其构造成分内层为果胶质(pectin),外层为硅质(
),使硅藻细胞壁成为坚硬的壳体。壳体由上、下两个半壳套合而成,像一个盒子。套在外面的较大的称为“上壳”,被套在里面的较小的称为“下壳”。每一个壳又由壳面、壳衣(壳套)和壳环三部分组成。壳面即硅藻细胞壁的顶面和底面,分别称作上壳面(盖板)和下壳面(底板),壳面平或呈凹凸状,辐射对称或两侧对称;壳衣(壳套)是壳面边缘略有倾斜的部分;壳环是与壳衣(壳套)相毗连,和壳面垂直的部分,上、下壳的壳环在自然套合的情况下总称连接带。有些硅藻在壳面和连接带之间还有附加的连接带,称为间生带,呈鳞片状或环状等。有些硅藻细胞壁上产生向细胞腔内延伸的板状突,称为隔片。间生带和隔片都具有增强细胞壁的作用。
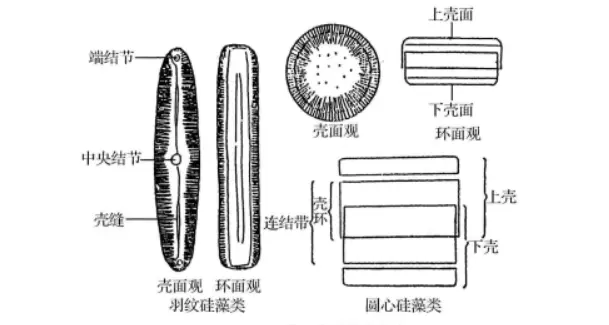

硅藻细胞的基本构造
硅藻细胞壁的壳面上具有排列规则的花纹,最常见的是由细胞壁上的许多小孔紧密或较稀疏地排列而成的线纹或点纹;有些种类在壳面内壁的两侧长有狭长横列的管状通道,形成“
”形的粗花纹,称为“肋纹”;有些种类在壳面边缘有纵走的凸起,称为“龙骨”。
花纹
花纹在硅藻细胞的壳面上,有排列细致而规则的各种花纹。较常见的花纹是由细胞壁上的许多小孔紧密或较稀疏排列而成的线纹。有的种类在壳面内壁的两侧长有狭长横裂的小室,形成呈“
”字形的粗花纹,称为“肋纹”。有的种类在壳的边缘有纵走的凸起,称为“龙骨”。壳面花纹的排列基本上为辐射排列与羽状排列二种类型。
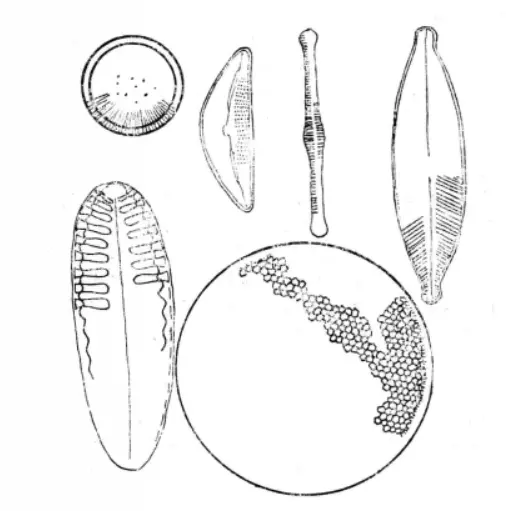
硅藻的各种花纹
壳缝
在羽纹硅藻纲的多数种类,于壳面中部或偏于一侧,有1条纵向的无纹平滑区,称为“中轴区”。沿中轴区中部或偏于一侧有一条纵向的裂缝与细胞内部相通,称为壳缝。壳缝一般呈直线形或“
”形、“
”形。
有的种类没有壳缝,仅有较窄的中轴区,称为假壳缝,有的种类的壳缝是一条纵走的或围绕壳缘的管沟,以极狭的裂缝与外界相通,称为管壳缝。管壳缝位于壳面边缘的龙骨凸起(称龙骨突)内,管沟的内壁具有数量不等的小孔与细胞内部相通,这些小孔称为“龙骨点”。壳缝与硅藻的运动有关,具壳缝和管壳缝的种类能运动,没有壳缝和具假壳缝的种类则不能运动。
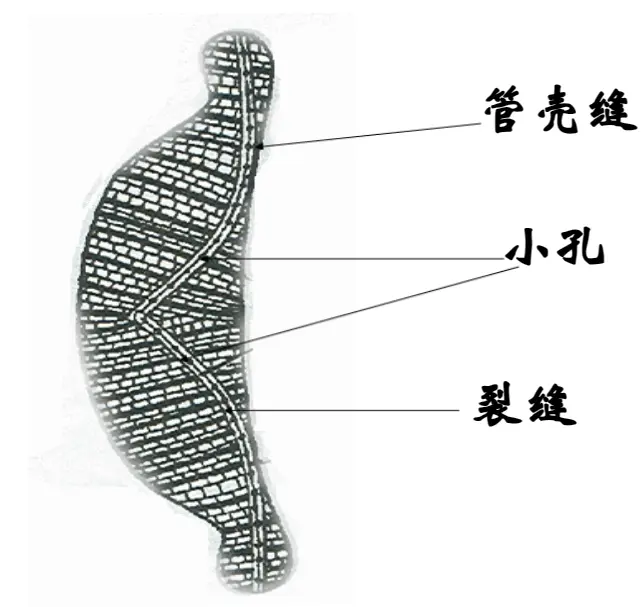
管壳缝
结节
在壳面中轴区中部,横线纹较短,形成面积稍大的“中心区”,中心区中部由于壳内壁增厚而形成中央结节。如壳内壁不增厚,仅具圆形或椭圆形或横矩形的无纹区,称为假中央结节。壳缝两端内壁各有一增厚部分,称为极(端)结节。
硅藻细胞的3个轴和3个面
3个轴分别是纵轴、横轴和贯壳轴。纵轴为壳面纵向的中轴线,又称顶轴、长轴;横轴为壳面横向的中轴线,又称切顶轴、短轴;贯壳轴为上下壳中心点相连线,又称壳环轴。这3个轴形成3个面,即壳面、长轴带面与短轴带面。在壳面为圆形的中心硅藻类,纵轴和横轴相等。
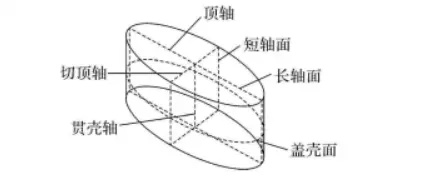
硅藻细胞的3个轴和3个面
色素和色素体
硅藻色素体中主要含有叶绿素
、
及胡萝卜素、硅甲藻素、岩藻黄素等,多呈黄绿或黄褐色。色素体一般贴近壳面,其形状及数目因种类不同而异。一般是中心硅藻纲的色素体多为小盘状,数量较多;羽纹硅藻纲的色素体多为大形的片状和星状,1个或2个,也有为多数小盘状的。
特征
编辑生态特征
硅藻种类很多,分布较广。几乎所有水体,如海洋、河口半咸水、江河、湖泊、水库、池塘甚至其他藻类难以繁殖的水体中都有硅藻的存在。在陆地上,凡是潮湿的地方,无论是土壤、墙壁、岩石及树皮上,以及在苔藓植物间都能生长,但主要分布在水域中。它们中的一些属、种只生活在淡水中;另一些属、种只能生活在海水中。即使是一些属所包含的物种既有海生的亦有在淡水中生活的,但是通常其中有一定的种绝对限于海洋,而另外有一定的种绝对限于淡水。在水体中,它们呈浮游状态或着生在其他物体上,着生种类常具胶质柄或包被在胶质团或胶质管中。
淡水藻类多生长在硬度较大的水中,密度有时会很大。条件适宜时一些营养丰富的水体会爆发硅藻水华。硅藻一年四季都能形成优势种群,一般在春秋两季大量繁殖。某些硅藻种类在水温高达60℃的温泉水中也可以正常生长繁殖。常见的硅藻水华优势种有直链藻、小环藻、冠盘藻、针杆藻和脆杆藻等。

尖针杆藻(Synedra acusvar)
生物学特征
硅藻为光合自养生物,依赖光和温度进行光合作用以快速生长。硅藻还存在与动物相类似的尿素循环,这种循环使硅藻能有效地利用碳和氮。尿素循环代谢产物是硅藻细胞碳循环和氮循环的关键,其代谢产物,对促进硅藻从营养缺乏状态中复苏并进行指数性增长十分重要和关键。
繁殖方式
编辑硅藻的繁殖主要有细胞有丝分裂和产生复大孢子或有性生殖。
细胞有丝分裂
细胞分裂时,原生质略膨大,璧的两半稍为分开,细胞核分裂成两个子核,载色体也随着分裂,分居细胞的一边。原生质体也沿着与瓣面平行的方向分裂为二,一个子原生质体位于母细胞上壳,另一个子原生质体位于母细胞的下壳。每个子原生质体各自再立即分泌出另一半细胞壁,新分泌出的另一半细胞壁,始终是作为子细胞的下壳。如此细胞每分裂一次,结果总是一个子细胞的体积和母细胞等大,一个子细胞必较母细胞更小。按照这样分裂下去,结果多数子细胞体积逐渐减小,当缩小到一定程度时,便产生复大孢子(auxospore)将细胞体积恢复到该种细胞的正常大小。硅藻的核分裂是在核内进行,纺锤体两端附着在核膜上,随着细胞及细胞核的伸长,染色体纺锤丝把染色体分别牵引向两端,分别形成子细胞核。
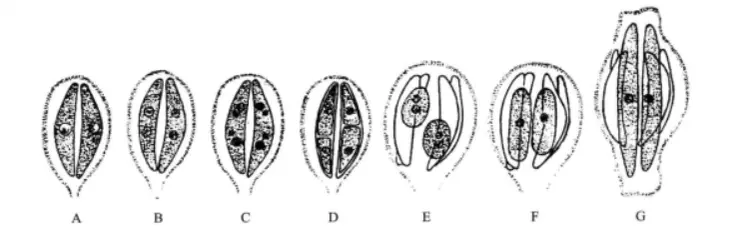
硅藻细胞分裂图解
复大孢子
复大孢子的产生为原生质体脱离两瓣,长大分泌新壁,成为复大孢子。复大孢子分裂产生大型的个体。在羽纹硅藻纲中,复大孢子由有性生殖产生。两个配偶的细胞接近,细胞核行减数分裂,两原生质体逸出即是配子,配子融合成合子合子长大成复大孢子。有时原生质体只分裂一次成两个配子,两细胞产生的4个配子结合成两个复大孢子。
有性生殖
通过异配、同配、卵配或自体受精等有性结合过程形成1个或2个合子。
常见种类
编辑硅藻门中常见的有直链藻属、根管藻属、桥弯藻属等。
直链藻属
截止2024年,直链藻属(学名:Melosira)公认有36个物种,多为浮游生物。该属细胞圆柱形,绝少数圆盘形、椭圆形或球形;壳面圆形,很少数为椭圆形,平或突起,有或无纹饰,有的带面常有1条线形的环状缢缩,称为“环沟”,环沟间平滑,其余部分平滑或具饰纹,有2条环沟时,两条环沟间的部分称为“颈部”;细胞间有沟状的缢入部,称为“假环沟”,壳面常有棘或刺;色素体小圆盘状。
直链藻种类在淡水、半威水和海水中均有分布,半咸水为主,河口区域最为常见。主要以淡水,早春、晚秋数量较多。在“水温”适宜的条件下,水体中可利用的“磷”含量增加,有助于直链藻“生长”,藻链长度增加,这也使直链藻加速沉降至“沉积物"表层,促进了水体中营养盐的清除,可以减缓高生产力“湖泊”中营养盐再生利用的效率。

直链藻
根管藻属
截止2024年,根管藻属(学名:Rhizosolenia)公认有65个物种,该属细胞直,扁圆柱形,或稍弯,常相连成长链,连接靠壳面的一个突起,或一个小刺,进入相连的细胞内。也有单独生活的。常见的面为壳环面,壳环面上的次级相连带(或称间板)呈鳞状或环状。突起或小刺的形态,也是重要的特征。色素体多,小盘状。大多浮游,海产,淡水产的很少,如距端根管藻。本属有休眠孢子,复大孢子和小孢子。

根管藻
桥弯藻属
桥弯藻属(学名:Cymbella),该属细胞单独生活,或以胶质柄附着在他物上,或在胶质管中生活。壳面纵轴两侧不对称,呈半月形、偏舟形或近舟形。壳缝偏向腹侧,直或弧状弯曲。具有明显的中轴区和中心区。点条纹通常放射状排列,点纹较为明显。有些种类,在中节向腹一侧,有特殊的眼纹状点纹。本属是典型的淡水种类,但也有少数出现在半咸水中。
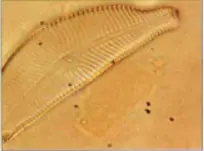
桥弯藻
双菱藻属
截止2024年,双菱藻属(学名:Surirella)公认有33个物种,该属藻体壳面线形、椭圆形或卵形,平直或呈螺旋状扭曲。壳面之两侧边缘具龙骨,龙骨上具管壳缝,管壳缝内壁具龙骨点。壳面有长或短的横肋纹,于肋纹间有纤细的横线纹。带面呈长形或楔形。色素体片状,1个。该属藻体常见,但出现量不大,常零星出现于各类水体中。
舟形藻属
截止2024年,舟形藻属(学名:Navicula)公认有408个物种,该属细胞三轴皆对称,单独生活,也有以胶质营、胶质块形成群体的。壳面多为舟形,也有椭圆形、菱形、棍棒形和长方形等。具壳缝、结节和由点组成的点条纹。每个细胞有载色体2~4个。该属生活在热带到寒带的海水、半咸水和淡水中,也很普遍地发现于化石里。在潮间带的经济贝类以及鱼类的消化道里都能大量找到。对于海水养殖业,作为人工培养饵料方面具有重要作用。
功用价值
编辑工业价值
硅藻的尸体沉积成为硅藻土,含有83%二氧化硅(SiO2),为工业用途极广的原料。由于硅藻土具有多孔性,容重小,其主要成分二氧化硅的化学稳定性好,耐热性能好,使其大量用作保温材料。硅藻土还有其他多种用途,如用作吸收剂、吸附剂、脱脂剂,过滤材料及水泥混合材料等。此外,硅藻可以小批量运输并从痕量增殖到理想的数量,且处理既不需要昂贵的材料也不需要复杂的仪器,是优良有效的工业原料。
生态价值
二氧化硅循环
硅藻在二氧化硅循环中起着重要的作用。世界大洋硅的生物地球化学循环系统包括五大分支系统,分别是可溶硅的输入、生物硅的生产、生物硅的埋藏、可溶硅的再循环和可溶硅的上涌。海洋硅藻作为最主要的生物硅生产者,直接或间接地参与了除硅输入之外的其他4个分支循环系统。从循环通量值来看,作为内部硅循环系统的生物硅生产、可溶硅再循环和可溶硅上涌参与的通量明显高于作为外部硅循环系统的可溶硅输入和生物硅埋藏,是海洋硅循环系统中最活跃的部分。另外,表层水体中硅藻生产的生物硅虽然含量很高,但只有不足3%的生物硅埋藏到海底,大部分生物硅因海水和孔隙水中可溶硅的不饱和而溶解,参与硅的再循环和上涌。
生物碳循环
硅藻是光合藻类,几乎遍布全球,是地球二氧化碳循环的重要组成部分,硅藻每年大约可以固定的二氧化碳量相当于全球总固定量的20%,其中海洋硅藻光合作用每年固定的碳与所有陆地雨林的总量相当,在生物地球化学循环中有至关重要的意义。硅藻在自身生长过程中利用光合作用不断吸收海水中的CO2,将无机碳转换成颗粒态有机碳。每年海洋中硅藻的有机碳净生产量比所有陆地热带雨林的有机碳净生产量还多。这些有机碳的大部分在微生物的作用下转化成无机态CO2,并在海洋中可以快速循环,少量有机碳可以快速沉降到海底,最终埋藏到海底沉积物中。单细胞硅藻体内生物化学机制的运行中最基本的代谢过程是在一系列酶的作用下,通过磷酸烯醇式丙酮酸、草酰乙酸和丙酮酸的相互转化。在单细胞内对CO2完成了收集、浓缩和转运等一系列过程,在叶绿体周围释放的CO2同RuBP发生反应生成3-PGA,可以有效降低光呼吸作用的反应强度,增加Rubisco的固碳效率。有些硅藻种类还涉及到了苹果酸丙氨酸和天冬氨酸等中间代谢物的相互转化。
天然饵料
硅藻占水生生物量的很大一部分,是重要的初级生产力,也是海洋食物网的重要组成部分。硅藻细胞内富含多种营养物,合成多种不饱和脂肪酸,是很多鱼类、贝类、虾类、鸟类和水生浮游动物的食物来源。因此,硅藻被认为是天然饵料的主要组成部分。在人工养殖经济生物时,硅藻常被大量培养作为养殖生物幼体时期的饵料。
研究价值
在水生生物生态学研究中,从20世纪早期开始直到现在长期被视为重要的生物指示类群用来监测水质和评价水环境。由于硅藻细胞壁在地层中能够保存,常被用来作石油勘探、地层划分和对比、古湖沼学的重建以及古地理、古气候方面的研究。此外,硅藻由于其外壳由多孔生物矿化硅组成,有着高度有序的层次结构,被认为是活着的“纳米结构工厂”,是纳米技术的理想材料。因此,学者们对硅藻纳米材料的研究已经进入各个领域,如染料敏化太阳能电池元件、生物传感器、药物运输载体、合成金属纳米粒子、电发光元件等。硅藻蛋白也因其独特的力学、光学、光激发性能和好的生物相容性而成为生物医药材料、纳米能源材料和环境功能材料等新兴科技材料。
其他用途
硅藻在商业应用中也有很大的价值,包括水产养殖业,生物活性物质,生物柴油等。如生物柴油是清洁的可再生能源,是优质的石油柴油代用品,但生物柴油产业受原料供给和成本的制约。而硅藻富含油(占细胞干重40%-60%),且短链脂肪酸(C14和C16)占细胞中总脂肪酸的67%-70%左右。其中,三角褐指藻细胞壁含硅较少,利于破碎,可作为优良的生物柴油生产原料。
危害
编辑硅藻门中的有些种类,如骨条藻(Skeletonema)、角毛藻(Chaetoceros)、海链藻(Thalassiosira)、拟菱形藻(Pseudo-nitzschia)的种类还是引发赤潮的原因种,有些还是有毒有害赤潮种类。例如拟菱形藻会形成软骨藻酸,人类和动物如果误食了被毒素污染的鱼类、贝类会出现急性中毒反应,严重者甚至死亡。此外,一些营附着生活的种类如曲壳藻、针杆藻、楔形藻等在紫菜采苗时,常大量附着在网线上,阻碍紫菜孢子的附着和小芽的生长,会影响紫菜生长。
相关研究
编辑有学者通过反复实验验证了硅藻的物理化学性质,利用动物对其溶血性、凝血性、细胞毒性、在体内体外的止血效果构建出血模型,以开发出不同的止血材料。证实了硅藻生物二氧化硅在止血方面的有效性,通过鼠尾截肢实验发现,硅藻的凝血时间远短于纱布的凝血时间。也有学者对硅藻的止血效果进行了研究,通过强酸洗涤纯化硅藻获得纯净外壳,其研究结果表明,硅藻具有优良的止血效果和良好的生物相容性及较低的细胞毒性。
此外,一些学者通过建立硅藻UPA条形码基因的实时荧光定量PCR检测方法,证实了基于硅藻UPA基因设计通用引物,建立的qPCR硅藻检验法应用于溺死诊断具有较好的应用前景。
参考资料
编辑展开[1]张彦文,周浓主编. 植物学[M]. 华中科技大学出版社, 2014.09: 70. (3)
[2]浏览分类树.物种2000(中国节点). [2024-02-29].
[3]叶庆华等编著. 植物生物学[M]. 厦门大学出版社, 2002.09: 285.
[4]莫雯惠. 基于细胞超微结构的肘杆藻属(硅藻门)分类研究[D]. 吉首大学, 2023 (4)
[5]袁莉. 基于细胞超微结构的脆杆藻科(硅藻门)分类研究[D]. 吉首大学, 2023 (7)
[6]魏洪祥. 水丰水库常见浮游生物图鉴及生态特征研究. 海洋出版社, 2022.08: 15. (3)
[7]无锡市环境监测中心站编著. 太湖常见藻类图集[M]. 中国环境出版社, 2017.09: 57.
[8]山东省水产学校主编. 淡水生物学 淡水渔业专业用[M]. 农业出版社, 1985.05: 33. (2)
[9]梁君荣教授团队揭示海洋硅藻休眠细胞形成的新机制.厦门大学生命科学学院官网. [2024-03-01].
[10]海洋硅藻存在与动物类似尿素循环
可使硅藻更有效地利用碳和氮.中国科学院. [2024-03-01].
[11]王毅超. 胶州湾硅藻生物多样性及其形成时间的超级条形码分析[D]. 中国科学院海洋研究所. 中国科学院大学, 2022
[12]王毅超. 胶州湾硅藻生物多样性及其形成时间的超级条形码分析[D]. 中国科学院海洋研究所. 中国科学院大学, 2022
[13]刘乾, 米铁柱, 甄毓, 等. 硅藻C4固碳途径的研究进展[J]. 海洋科学, 2018
[14]王雨璇, 陈冠虹, 喻敏, DE Vadim, 李芳柏, 方利平. 淡水硅藻的砷甲基化和砷氧化代谢机制[J]. 环境工程学报, 2023
[15]李铁刚,熊志方,翟滨著. 低纬度西太平洋硅藻席沉积与碳循环[M]. 海洋出版社, 2015.08: 4.
[16]湖南省水产科学研究所编. 淡水渔业实用手册[M]. 湖南科学技术出版社, 1984.04: 275.
[17]李红霞主编;徐延庆,王守业,柴俊兰,王战民等副主编. 耐火材料手册 第2版[M]. 冶金工业出版社, 2021.01: 597.
[18]海藻学. 海藻学[M]. 中国海洋大学出版社, 2014.05: 470.
[19]齐雨藻等著. 中国南海赤潮研究[M]. 广东经济出版社, 2008.12: 69.
[20]李扬汉著. 植物学 中[M]. 高等教育出版社, 1987.10: 121.
[21]龙继艳. 基于细胞超微结构的双菱藻科(硅藻门)分类研究[D]. 吉首大学, 2023 (7)
[22]金德祥著. 海洋硅藻学[M]. 厦门大学出版社, 1991.01: 75. (11)
[23]齐雨藻, 李家英, 高亚辉, 等. 硅藻分类系统与系统学研究进展[C]. 第二届全国藻类多样性和藻类分类学术研讨会论文集.2010, 2010: 22-23. (2)
[24]Bacillariophyceae.ITIS. [2024-03-01].
[25]周阳艳. 基于超微结构的桥弯藻属(硅藻门)分类研究[D]. 吉首大学, 2022 (3)
[26]石彭灵,黄洋主编. 水生生物学[M]. 电子科技大学出版社, 2020.04: 44. (2)
[27]钱树本主编. 海藻学[M]. 中国海洋大学出版社, 2014.05: 468.
[28]马放,杨基先,魏利等著. 环境微生物图谱[M]. 中国环境科学出版社, 2010.06: 45. (6)
[29]Melosira.World Register of Marine Species. [2024-03-12].
[30]王超, 刘杨, 李新辉, 等. 基于文献计量学的直链藻属的研究进展[J]. 生态学报, 2016
[31]Rhizosolenia.World Register of Marine species. [2024-03-20].
[32]Surirella .World Register of Marine species. [2024-03-20].
[33]山东省水产学校主编. 海洋浮游生物学[M]. 中国农业出版社, 1993.05: 8. (2)
[34]Navicula.World Register of Marine species. [2024-03-20].
[35]刘晓丹, 任庆敏, 王寅初, 等. 硅藻纳米材料研究进展[J]. 生物学杂志, 2019
[36]王妃. 东海和黄海硅藻、颗粒物及沉积物中的甾醇类生物标志物比较研究[D]. 中国海洋大学, 2013 (2)
[37]陈春艳, 胡晗华, 王煜, 等. 氯化铁和聚丙烯酰胺絮凝剂WT652对三角褐指藻的絮凝作用[J]. 水生生物学报, 2015
[38]宋蕴琦, 王寅初, 王绪敏. 硅藻纳米止血材料的研究进展[J]. 黑龙江科学, 8646 (2)
[39]刘向东, 刘超, 徐曲毅, 等. 实时荧光定量PCR检测硅藻UPA基因在溺死诊断中的研究[J]. 中国法医学杂志, 5728
内容由百科小编提供,本内容不代表globalbaike.com立场,内容投诉举报请联系全球百科客服。如若转载,请注明出处:https://globalbaike.com/27942/