- 1 定义
- 2 基本特征
- ▪ 数量特征
- ▪ 空间特征
- ▪ 遗传特征
- ▪ 系统特征
- 3 空间格局
- ▪ 随机分布
- ▪ 均匀分布
- ▪ 集群分布
- 4 种群数量与动态
- ▪ 种群大小和密度
- ▪ 样带抽样法
- ▪ 样方抽样法
- ▪ 标志重捕法
- ▪ 自然种群的数量变动
- ▪ 季节消长
- ▪ 年变化
- ▪ 周期性波动
- ▪ 不规则波动
- ▪ 种群爆发
- ▪ 人为干扰下的种群动态
- ▪ 原生环境的破坏对种群动态的影响
- ▪ 生物资源的过度猎取
- ▪ 环境污染后的种群动态
- 5 种群数量变动影响因素
- ▪ 出生率
- ▪ 死亡率
- ▪ 迁入率和迁出率
- ▪ 其他作用因素
- ▪ 性别比例
- ▪ 年龄结构
- ▪ 动物行为
- 6 种群动态描述
- ▪ 动态模型
- ▪ “J”型增长
- ▪ “S”型增长
- ▪ 生命表
种群(population)是一种生物学概念,指的是特定时间内占据一定空间的同种生物的所有个体。种群是群落的基本组成部分和生态学研究的最小生态单位,具有数量特征、空间特征、遗传特征和系统特征4大基本特征。其中数量特征是种群的最基本特征,受出生率、死亡率、迁入率和迁出率、种群的年龄结构和性别比率等因素影响;空间特征即其个体在空间上的分布格局,可分为集群分布、随机分布和均匀分布。
定义
编辑种群(population)是特定时间内占据一定空间的同种生物所有个体的总称。种群与物种概念密切相关,根据生物学对于物种的定义,同一物种的个体不仅因其同源共祖而表现出性状上的相似,且它们之间能相互交配并将其性状遗传给后代个体。不同种之间则由于形态、生理或行为上的差异而不能交配繁育,这种现象称为生殖隔离。与此对应,广义的种群即是指一切可以交配并产生可育后代的同种个体集群,即该物种的全部个体。
种群是物种的存在单位、繁殖单位和进化单位,也是群落的基本组成部分和生态学研究的最小生态单位。在生物组织层次结构中,种群代表由个体水平进入群体水平的第一个层次。由于有性生殖过程是一个基因重组过程,重组产生新的变异,可供自然选择,所以相互交配繁育的种群便构成了一个进化单位,它可能成为分化新物种的起点,有的生物还由繁育关系组成一定的社群结构。另一方面,同一地区的同种个体共享同一资源,因而又表现出种内竞争或合作关系。
基本特征
编辑种群具有数量特征、空间特征、遗传特征和系统特征4大基本特征。
数量特征
种群的数量特征是种群的最基本特征。种群是由多个个体组成的,其数量大小受到出生率、死亡率、迁入率和迁出率这四个种群参数影响。此外,种群的年龄结构、性别比率等也会影响种群的数量。
空间特征
空间特征即其个体在空间上的分布格局。受到种内关系、种间关系及自然环境条件的综合影响,种群在空间分布上呈现出随机分布、均匀分布和集群分布等不同的空间分布格局。
遗传特征
种群是同种个体的集合,因此具有一定的遗传组成,种群中全部个体所含的全部基因构成了这个种群的基因库。不同的地理种群之间存在有基因差异,也具有不同的基因库。种群的基因频率世代传递,在进化过程中通过改变基因频率以适应环境的不断改变,从而决定了生物进化的方向。
系统特征
种群是一个自组织、自调节的系统,是以一个特定的生物种群为中心,以作用于该种群的全部环境因子为空间边界组成的系统。因此,从系统角度研究种群内在因素及生境内各种环境因素与种群数量变化之间的相互关系,可揭示种群数量变化的机制与规律。
空间格局
编辑组成种群的个体在其空间中的位置状态或布局,被称为种群空间格局(spatial pattern)或内分布型(internal distribution pattern)。种群的空间格局可分为3类:随机分布、均匀分布、集群分布。
随机分布
当个体之间的相互作用并不强烈或者它们的生活环境不均匀时,种群中的个体就会呈现随机分布的形式。随机分布比较少见,因为在环境资源分布均匀,种群内个体间没有彼此吸引或排斥的情况下,才易产生随机分布,如森林地被层中的一些蜘蛛(Araneida),面粉中的黄粉虫(Tenebrio molitor)等。
均匀分布
均匀分布指的是种群在空间按一定间距均匀分布产生的空间格局,在自然界中可能较为常见。但由于个体之间对于资源的竞争,因此在种群中也并不总是呈现均匀分布的形式。这就意味着它们通过竞争来实现这一分布形式,且方式各不相同。如在动物界中,均匀分布常常是由动物之间行为上的相互作用引起的。在许多物种中,某一性别或者两种性别的物种通过将其他的个体排除在外来捍卫自己的领域。这些领域专一地为占领者提供各种资源,如食物、水、避难空间以及配偶等,因而个体往往均匀地分布在它们所占有的栖息地上。甚至在一些没有自己领域的物种中,它们也常常会维护一片不允许其他动物侵入的防御空间。
在植物界,均匀分布也是由于个体间竞争资源产生的一种常见现象。聚集在一起的植物个体将会竞争阳光、营养成分以及水等。这种竞争可能是直接的,比如一个植物体将另一个植物体遮挡起来。这种竞争也可能是间接的,比如两个植物体通过吸收同一片区域内的营养成分或是水产生竞争。此外,有些植物,如三齿团香木(Larrea tridentata),能向周围土壤中分泌化学物质,这种物质对其他的物种个体是有害的,在这些情况下,植物个体之间只有保持足够的距离才能实现共存,从而导致均匀分布。
集群分布
个体聚集成群是对环境中资源分布不均的响应,集群分布在自然界中较为普遍。动植物个体以及微生物更喜欢根据土壤类型、湿度以及其他它们最能适应的环境特征来划分的小生境。此外,社交互动也可以导致集群分布。许多物种在一个大型的群体中生活并四处移动,这种群体具有很多优势、比如提高防御捕食者的意识、降低能量消耗,也可以获取所有群体成员的信息。
在更广泛的范围内,种群常常在活动领域范围内密集地分布,靠近边界的种群分布较为稀少,这种分布模式是由不同区域内的环境变化引起的。种群常常最能适应它们自身分布领域内的环境条件,随着环境状况发生改变,个体适应能力下降,因此种群密度随之下降。
种群的分布类型
种群数量与动态
编辑种群大小和密度
种群大小指的是一个种群中的个体数量,单位面积或容积内的个体数目被称为种群密度。种群密度是影响种群数量动态的重要因素之一,也是种群的一个重要参数。种群密度可以用单位面积或空间内的个体数目来表示,也可以用生物量或能量来表示,这是因为不同的生物种其个体大小、代谢率均不同。在食物链中营养层次越低,密度越高,如一些小型生物。在这种情况下,能量流动是衡量生态系统任何种群或全部生物种群较为合适的指标。
种群密度可分为绝对密度(absolute density)和相对密度(relative denaity),前者指单位面积或空间上的个体数目,后者是表示个体数量多少的相对指标。绝对密度测定分为总数量调查法和取样调查法。总数量调查法是计数某面积内全部生活的某种生物的数量,如一个保护区内大熊猫的数量。取样调查法是指在总数量调查比较困难的情况下所采用的一种方法,它只计数种群中的一小部分,用以估计整体。该调查方法包括样带抽样法、样方抽样法、标志重捕法等。
样带抽样法
样带抽样法适宜动物调查,其基本原理是调查者沿着一条预先设定的路线行走,通过记录样带中所见到和听到的动物个体数或动物痕迹数,取得种群绝对密度或相对密度的估计值。
样方抽样法
样方抽样法适合调查植物以及活动能力不强的动物,如蜗牛(Fruticicolidae)、蒲公英(Taraxacum mongolicum)等。具体操作过程是:在被调查范围内,随机选取若干个完全相等的样方,统计每个样方的个体数,并求出每个样方的种群密度,再求出所有样方种群密度的均值,以此值作为被调查种群之种群密度的估算值。计算公式为D(密度估计值)=N(抽样面积上计数的个体总数)/A(抽样面积)。常见的取样方法有等距取样法、五点取样法、Z字取样法等。
标志重捕法
标志重捕法适用于活动能力较强、活动范围较大的动物,如田鼠(Arvicolinae)、鸟类(Aves)等。具体操作过程是:在被调查种群的活动范围内,捕获部分个体,作上标记,再放回原来的环境中,经过一段时间后在同一地点进行重捕。估算公式为M(首次捕获并标志的个体数)/N(种群大小)=R(第二次捕获的个体中已被标志的个体数)/n(第二次捕获的个体数),N(种群大小)=M(首次捕获并标志的个体数)n(第二次捕获的个体数)/R(第二次捕获的个体中已被标志的个体数)种群总数/标记个体数=重捕个数/重捕中标记个体数。
此估算方法得出的估算值偏大,因为很多动物在被捕获一次后会更加难以捕获,导致重捕中标记个体数偏小。标记时也需要注意,所用标志要小而轻,不能影响生物行动,也不能用过于醒目的颜色,如红色,否则会使生物更加容易被天敌捕食,影响估算精确度。
自然种群的数量变动
自然界种群的数量变动,除了一般的增长和消亡外,比较明显的是季节消长和年变化。自然种群的研究表明,种群数量具有两个重要特征:1. 波动性,即在每一时间段(年、季),种群数量都有所不同;2.稳定性,尽管种群数量存在一定的波动,但大部分的种群不会无限制地增长或无限制地下降而发生灭绝,也就是说虽然种群中有出生和死亡,其成员也在不断更新,但某种程度上这种变动都往往维持在特定的水平上。种群数量在相当长的时期内维持在一个水平的状态称为种群平衡,这种平衡是相对的,指的是一年中出生数和死亡数大致相当,种群数量基本稳定。
季节消长
种群季节消长是指种群数量在一年四季中的变化规律,种群数量消长规律是种群数量动态规律之一。一般具有季节性生殖的种类,种群的最高数量常落在一年中最后一次繁殖之末,以后其繁殖停止,种群因只有死亡而无生殖,故种群数量下降,直到下一年繁殖开始,这时是种群数量最低的时期。季节消长常出现在体型较大、一年只繁殖一次的动物,如狗獾(Meles meles)、旱獭(Marmota bobak)等,其繁殖期在春季,产仔后数量达到高峰,以后由于死亡,数量逐渐降低。对这类动物的数量调查,通常要进行两次。
年变化
种群数量在不同年份的变化有的具有规律性(周期性),有的无规律性。种群动态研究表明,大多数种群的年变化表现为不规律的波动,有周期性数量变动的种类是有限的。在环境相对稳定的条件下,种子植物及大型脊椎动物的种群密度在较长的时间跨度内呈现周期性变化,如杨(Populus)、柳(Salix)等乔木每年开花结果一次,种子数量相对稳定。资料表明,中国黑龙江伊春林区的小型鼠类种群也表现出3-4年的波动周期,每遇高峰年的冬季就会造成林木危害,尤其是幼林。
周期性波动
周期性波动(regular fluctuation)是指种群数量在不同的时间进程中表现出规律性或周期性变动的现象。在一些情况下,捕食或食草作用造成的密度减小会导致种群出现周期性波动。如生活在瑞士森林中的灰线小卷蛾(Zeira phera griseana),在春天随着落叶松的生长,其幼虫也会同时出现,而幼虫的吞食会减小松针大小,从而使翌年幼虫的食物质量下降,最后导致灰线小卷蛾种群数量下降。反之,低密度的幼虫数量使落叶松松针大小恢复,但随着食物质量提高,幼虫数量又开始增加。
不规则波动
环境的随机变化容易造成种群的不规则波动,有一些动物的种群数量年变化很剧烈,但不具有周期性,如小家鼠(Mus musculus),据中国科学院的2016年的统计资料,其年均捕获率波动于0.10-17.57之间。布氏田鼠(Lasiopodomys brandtii)也具有不规律的数量变动,其数量最低的年代平均每公顷只有1.3只鼠,而数量最高的年份每公顷可达786只。但是这种不规律的变动都会围绕着一个平均密度,即种群受某种干扰而发生数量的上升或下降,有重新回到原水平的倾向,这种情况就是动态平衡。
种群爆发
具有不规则或周期性波动的生物都有可能出现种群爆发,如赤潮,即水中的一些浮游生物爆发性增殖,从而引起水色异常。赤潮主要发生在近海,是水中氮、磷等营养物过多形成富营养化所致,主要危害有藻类死体分解,消耗大量的水中溶解氧,使鱼贝等窒息而死。此外,飞蝗和鼠类也是最常见的易爆发物种。
赤潮
人为干扰下的种群动态
人为干扰指的是有毒化学物质的释放、森林砍伐、筑坝、草原开垦、过度放牧和开垦耕地等人类活动。
原生环境的破坏对种群动态的影响
原生环境储藏着自然资源,具有生态实验室、遗传库和信息储存库的作用,野生动物种群可使这种自然系统获得平衡。在人类过度捕猎或栖息地被破坏的情况下,种群长期处于不利条件下,其数量可能出现长期下降,甚至出现种群绝灭。物种的自然灭绝的速度是很缓慢的,大约每千年有一种灭绝,而人类活动会使野生动物的灭绝速度加快。种群濒危和灭绝的原因主要是原生环境遭到破坏,居住空间逐渐缩小,使其无地容身,因此,野生生物的保护越来越集中在保护栖息地方面。
森林砍伐
生物资源的过度猎取
种群长期处于过度猎捕的条件下,其种群数量将下降或灭亡。如1913年,太平洋鲑鱼(Salmo salar)的产量为240万箱,当年在鲑鱼主要产卵河流上修筑了铁路,第一次破坏了产卵周期,其捕获量立即开始下降,到1923年,捕获量仅有9万箱。
捕获太平洋鲑鱼
环境污染后的种群动态
环境污染会导致种类数量减少,只有少数种类或个别种类的数量增加。原有生物与环境中各种物质关系发生变化,出现了新的生物与环境的物质循环关系,一般是耐污种类在污染环境中增多,而耐污性较弱的种类消失,狭污性种类被广泛性种类所代替。
种群数量变动影响因素
编辑种群的数量会随着时间而变化,因此需考虑反映数量变化的各种因子,包括死亡率、出生率、迁入率和迁出率。个体的出生和迁入使种群数量增加,死亡和迁出使种群数量减少。如果增量大于减量,种群数量增加,反之减少;如果增量与减量相等,则维持不变。
出生率
出生率是指种群产生新个体占个体数的比率,常以单位时间内产生的新个体数来表示,有绝对出生率和相对出生率两种表示方式。绝对出生率B(新产生的个体数)= ΔN(新产生的个体数)/Δt(时间增量),相对出生率B(新产生的个体数)= ΔN(新产生的个体数)/[N(种群的总个体数×Δt(时间增量)]。
出生率可用生理出生率和生态出生率来表示。生理出生率又叫最大出生率,是种群在理想条件下(即无任何生态因子的限制作用,只受生理因素所限制)所能达到的最大出生率,对某个特定的种群,是一个常数。生态出生率又叫实际出生率,是指在某个真实的或特定的大环境下种群的实际出生率,这是在自然条件下经常出现的出生率,不是固定的,是随着种群大小、组成和物理环境条件的不同而变化。
死亡率
死亡率是用来描述种群中个体死亡的速度,是种群内数量衰退的因素。死亡率也可以用生理死亡率(可最小死亡率)和生态死亡率(实际死亡率)来表示。生理死亡率是种群的常数,表示在理想的或无限制的条件下死亡的个体数。种群的生理寿命是指种群处于最适条件下的平均年龄,而不是某个特殊个体可能具有的最大寿命。生态死亡率是在自然条件下真正的死亡率,它也像生态出生率一样不是一个常数,是随着种群状况和环境条件而改变,即种群内的个体达到生态寿命长度。种群的生态寿命是指种群在特定环境条件下的平均寿命。死亡率的大小与生物的遗传特性、年龄组成和生活环境有密切关系。
迁入率和迁出率
许多生物种群存在着迁入、迁出的现象,大量个体的迁入或迁出会对种群密产生显著影响。对于一个确定的种群,单位时间内迁入或迁出种群的个体数占种群个体总数的比例,分别成为种群迁入率(immigration rate)或迁出率(emigration rate)。有些自然种群持久的迁出个体,保持迁出率大于迁入率,有些种群只依靠不断的迁入才能维持下去。植物种群中迁出和迁入的现象相当普遍,如孢子植物借助风力把孢子长距离地扩散,不断扩大自己的分布区。种子植物借助风、昆虫、水及动物等因子,传播其种子和花粉,在种群间进行基因交流,防止近亲繁殖,使种群生殖能力增强。
其他作用因素
除死亡率、出生率、迁入率和迁出率外,种群结构本身的特点如性别比例、年龄结构、动物行为等,也能影响种群数量。
性别比例
种群的性别比例(Sex Ratio)或性别结构(Sexual Structure)指的是种群中雄性个体与雌性个体的比例,通常用每100个雌性的雄性数来表示,即以雌性个体数为100,计算雄性与雌性的比例。如果性比例等于1,表示雌雄个体数相等;如果大于1,表示雄性多于雌性;如果小于1,表示雄性少于雌性。
自然界中,不同种群的正常性别比例有很大差异,性别比例对种群数量有一定影响。如人(Homo sapiens)、猿(Anthropoidex)等高等动物的性比例为1,鸭科(Anatidae)、一些鸟类及许多昆虫的性比例大于1,蜜蜂(Apoidea)、蚂蚁(Formicidae)等社会昆虫的比例小于1。此外,种群的性别比例会随着个体发育阶段的变化而发生改变,如啮齿类(Glires)动物出生时性别比例为1,但3周后的性别比例为1.4。性比例影响着种群的出生率,因此也是种群数量变动的因素之一,对于一雄一雌婚配的动物,种群当中的性比例如果不是1,就必然有一部分成熟个体找不到配偶,从而降低了种群的繁殖力,对于一雄多雌、一雌多雄分配制,以及没有固定配偶而随机交配的动物,一般来说,种群中雌性个体的数量适当地多于雄性个体有利于提高生殖力。
年龄结构
种群的年龄结构(Age Structure)就是不同年龄组(Age Classes)在种群中所占比例或配置状况,其对种群出生率和死亡率均具有较大影响。多采用年龄锥体(Age Pyramid)或年龄金字塔来分析年龄结构,年龄金字塔是从下到上的一系列不同宽度的横柱做成的图。横柱高低位置表示从幼年到老年的不同年龄组,横柱的宽度表示各个年龄组的个体数或其所占的百分比。按美国学者博登海默(Bodenheimer)1958年的划分法,年龄金字塔可分为以下3个基本类型:
增长型种群(Expanding Population):左侧A为锥体结构,具有宽的基部,而顶部狭窄,表示幼体的百分比很高,就是说种群中有大量的幼体。而老年的个体却很少,这样的种群出生率大于死亡率,是迅速增长的种群。
稳定型种群(Stable Population):中间B为钟型结构,说明种群中幼年个体和中老年个体数量大致相等,其出生率和死亡率也大致平衡,种群数量稳定。
下降型种群(Diminishing Population):右侧C锥体呈壶型,基部比较窄而顶部比较宽,表示幼体所占的比例很小,而老年个体的比例较大,种群死亡率大于出生率,是一种数量趋于下降的种群。
A.增长型种群,B.稳定型种群,C.下降型种群
动物行为
动物的行为如扩散、聚集与迁移等均可间接影响种群数量。如扩散使种群密度下降,疾病传染、食物竞争、生殖力降低等对种群密度有影响的控制因素无法起到作用,种群得以继续增长。聚集则会使种群密度增加,对密度有影响的控制因素能够发挥作用,抑制种群进一步的增长,甚至导致死亡率上升。
种群动态描述
编辑动态模型
种群中一些简单的、具有典型性的动态变化可以用模型衡量,常见的为以下2种:
“J”型增长
“J”型增长为理想状态下的种群增长,即种群处于空间、食物等资源无限的环境中,因而其增长率不随种群本身密度的变化而变化。这类增长通常呈现指数增长形式,且与密度无关。与密度无关的增长又可分为两类:1. 种群的各个世代彼此不相重叠,如一年生植物和许多一年繁殖一次的昆虫,其增长是不连续且分步的,因此被称为离散增长,一般用差分方程描述。2. 如果种群的各个世代彼此重叠,如人类和多数兽类,其种群增长是连续的,可用微分方程描述。
“S”型增长
在自然界中,环境条件是有限的。因此,种群不可能按照“J”形曲线无限增长。当种群在一个有限的环境中增长时,随着种群密度的上升,个体间对有限的空间、食物和其他生活条件的种内竞争必将加剧,以该种群生物为食的捕食者的数量也会增加,致使这个种群的出生率降低,死亡率增高,从而使种群数量的增长率下降。当种群数量达到环境条件所允许的最大值(以K表示)时,种群数量将停止增长,有时会在K值附近保持相对稳定。假定种群的增长率随着种群密度的增加而按一定的比例下降,种群数量达到K值后保持稳定,那么种群的增长就会呈出“S”形曲线。
种群增长曲线
生命表
生命表(life table)又被称为死亡表,是描述种群动态发展过程的重要方式。生命表是按种群生长时间或年龄(发育阶段)的次序编制的,可系统记述种群死亡率、生存率和生殖率,直观展示种群死亡和存活的过程。生命表可提供种群基本信息,计算种群统计值,绘制存活曲线,同时能进行关键因子分析。生命表可分为动态生命表、静态生命表和综合生命表。
动态生命表
动态生命表又叫同生群生命表,是同一时间段出生的个体从出生到死亡的数据统计,即同时出生/孵化的一个群体(同生群),跟踪观察并记录其死亡过程直到最后。
静态生命表
静态生命表是根据在某一特定时间对种群进行年龄结构的调查所得数据编制的,一般在难以获得动态生命表的情况下使用。表中数据是根据在某一特定时刻对种群年龄分布频率的取样分析而获得的,实际反映了种群在某一特定时刻的剖面。
综合生命表
动态生命表个体经历了同样的环境条件,而静态生命表中个体出生于不同的年份,经历了不同的环境条件。因此,编制静态生命表等于假定种群所经历的环境没有变化。事实上情况并非如此,所以有的学者对静态生命表持怀疑态度。但动态生命表有时历时长,工作量大,往往难以获得生命表数据。静态生命表虽有缺陷,在运用得法的情况下还是具有一定参考意义的。因此,一般世代重叠且寿命较长的生物如人类,宜编制静态生命表。而对于世代不重叠的、生活史比较短的生物如某些昆虫,则宜编制动态生命表。
与动态生命表和静态生命表相比,综合生命表增加了描述种群各年龄出生率的指标,指的是同种群平均每存活个体在该年龄期内所产生的后代数。
存活曲线
存活曲线(survival curves)指的是特定年龄存活个体占初始种群数量的百分比,可直观描述种群发展的动态过程。存活曲线以生物的相对年龄(绝对年龄/平均寿命)为横坐标,以各年龄的存活数的对数为纵坐标。存活曲线是表达年龄分布的重要途径,主要分为以下3种类型:
A型:曲线凸型。种群在达到生理寿命之前只有少数个体死亡,只在生命末期死亡率才较高,人类和一些大型哺乳动物的存活曲线属于此类。
B型:对角线型。各年龄段的死亡率基本相等,如蝎蝽(Nepidae )、鸟类等。
存活曲线的类型
自然增长率
在自然界中,种群的实际增长率称为自然增长率(rate of natural increase),用r来表示,指的是单位时间内某一种群的增长百分比。种群增长率可由生命表计算得出,一般生命表中都有存活率(lx),在生命表中加入一项特定年龄出生率(mx),就构成了综合生命表。把各年龄组的lx与mx相乘,并累加起来,可以得到净增殖率(R0),具体公式为R0=∑lxmx,。如当R0=3.1时,表示该生物经一个世代将增长到原来的3.1倍。种群增长率r=InR0/T,其中T为平均世代长度,指种群中个体从母体出生到其产子的平均时间。从公式中也可以看出,种群增长率r随R0增大而变大,随T值增大而变小。
内禀增长率
种群增长率r随着自然界环境条件的改变而发生变化。当条件有利时,r值可能是正值,种群增加;条件不利时可能变为负值,种群数量下降。如在实验室条件下,排除不利的天气条件及捕食者和疾病等不利因素,提供理想的食物条件,就可以观察到种群的最大增长能力,研究人员将其称为种群内禀增长率,用rm表示。1954年,澳大利亚生态学家安德烈沃斯(Andrewartha)和伯奇(Brich)对种群的内禀增长率给出了明确定义,即具有稳定年龄结构的种群,在食物与空间不受限制,密度维持在最适水平,环境中没有天敌,并在某一特定的温度、湿度、光照和食物性质的生境条件组配下,种群达到的最大增长率。由定义可以看出,人们只能在实验室条件下才能测定种群的内禀增长率。rm可以作为一个参数,与自然界中观察到的实际增长率进行比较。
种群调节
编辑在自然界中,绝大多数生物的种群都处于一个相对稳定的状态,由于生态因子的作用,使种群在生物群落中与其他生物成比例地维持在某一特定密度水平上,这种现象被称为种群的自然平衡,此时的密度水平称为平衡密度,种群离开平衡密度又回到这一平衡密度的过程称为种群的调节。种群的数量变动是种群出生率和死亡率的变化过程,以及迁人、迁出的相互作用的综合结果。自然界中决定种群数量变化的因素是多种多样的,生态学家提出了很多不同的学说来解释种群的动态,可概括为两大类,即内源性因子和外源性因子。
制约因素
自然种群受到各种因素的制约,其中密度因素是调节种群平衡的重要因素。种群调节的因素,可分为密度制约(density dependent)和非密度制约(density independent)两类。密度制约因素是与种群本身的密度有关的作用因素,种群的死亡率随着种群密度的增加而增加,如种间竞争、捕食者、寄生以及种内调节等生物因素都属于密度制约因素。非密度制约因素是与种群本身的密度无关的作用因素,种群的死亡率不随种群密度的变化而变化,非密度制约因素包括天气条件、污染物以及其他环境的理化等非生物因素,这些因素有时会影响种群的数量,甚至可以使生物绝灭。
密度调节
密度调节是指通过密度因子对种群大小的调节过程,主要包括种内调节和种间调节两个部分。
种内调节
种内调节强调内源性因素的作用,指的是种内成员之间因行为、生理和遗传的差异而产生的一种密度制约性调节方式。主张自动调节的学者的研究重点在于动物种群内部,其主要观点包括:1. 强调种群内成员表现在行为、生理特征或遗传特征上的异质性;2. 种群密度的变化可影响种内成员;3. 将种群调节看成物种的一种适应性反映,经过自然选择后可带来进化上的利益。自我调节理论学者重视进化方面的论据,其认为种群波动的过程为种群增长→种内变异增加→很多劣质基因被保存下来→条件恢复正常→自然选择淘汰劣质个体→种群数量下降。自动调节假说可分为行为调节、内分泌调节和遗传调节。
1962年,英国生态学家温恩·爱德华兹(Wynne Edwards)通过对鸟类的研究提出行为调节假说。他认为动物通过社群行为可以限制其在生境中的数量,使食物供应和繁殖场所在种群内得到合理分配。当种群密度超过一定限度时,领域的占领者会产生抵抗,把多余的个体从适宜的生境中排挤出去,这部分个体由于缺乏食物以及保护条件,易受捕食、疾病、不良天气所侵害,死亡率较高,从而限制了种群增长。
内分泌调节又称生理调节,指的是种内个体间因生理功能的差异,生物功能强的个体在种内竞争中取胜,淘汰弱者。1950年,美国学者克里斯琴(Christian)提出内分泌学说,克里斯琴认为当种群数量上升时,种内个体间的社群压力增加,个体间处于紧张状态,加强了对中枢神经系统的刺激,影响脑下垂体和肾上腺的功能,使生长激素减少、生长和代谢发生障碍、生殖受到抑制,从而导致胚胎死亡率增加,出生率降低,种群数量下降。种群密度低时,社群压力降低,通过生理调节又可以恢复种群数量。
1960年,美国学者奇帝(Chitty)提出遗传调节假说,他认为种群中的遗传双态或遗传多态现象具有调节种群的作用。如种群中有两种遗传型,一种是繁殖力低、进攻性强、适于高密度条件下的基因型A,另一种是繁殖力高、适于低密度条件的基因型B,当种群数量较低时,自然选择有利于基因型B的个体,能相互容忍,繁殖力高,种群数量增加;当种群数量上升到较高水平时,自然选择转向对基因型A的个体有利,个体之间相互进攻,死亡增加,生殖减少,于是种群数量下降。因此,任何改变出生率和死亡率的因素都会影响种群的平衡密度,从而调节种群数量。
种间调节
种间调节是密度调节内容之一,指的是捕食、寄生和种间竞争共同资源因子对种群密度的制约过程。强调外源性因子的作用,主张这些生物因素对种群密度调节起决定作用的是生物学派。生物学派认为动物种群在正常情况下处于平衡之中,并在有限的范围内活动。因此,种群是一个自我管理系统,必然存在着一个平衡密度,而这一密度是由生物的捕食、寄生、竞争等密度制约因子来调节的。
非密度调节
非密度调节是指非生物因子对种群大小的调节,其也强调外源性因子的作用。气候因子对种群影响很大,最早提出气候因素对种群密度有调节作用是美国学者博登海默(Bodenheimer),他认为天气可通过影响生物的发育与存活来决定其种群的密度。气候学派主张种群密度主要靠气候等非密度制约因子来调节,强调波动性,不太重视稳定平衡。早期气候学派的主要观点是种群参数受气候因子的强烈影响,种群的密度波动发生与气象因子显著相关,强调种群的波动,否认种群的稳定。
后期气候学派的主要观点是反对区别生物因子和非生物因子,如植物,既可作为植食性动物的食物(即生物因子),也可作为庇护所(即非生物因子),为取食它的生物提供避难场所,反对将环境因子与种群密度直接联系在一起,认为当种群密度较大时,会占据更多的边缘生境。一些非生物因子(如严寒)的作用却与种群密度相关,这些因子对大种群的影响较大,而对小种群的影响较小,将动物种群生态因子分为气象、食物、其他动物、栖息地,它们都能对种群的数量和分布起决定作用。
内外因相结合
随着现代生态学的发展,种群调节机制研究的方法也逐渐增多,如系统生态学的引入,将横向思维代替了纵向思维,研究人员认识到利用单一的内因或外因来解释种群的调节机制过于简单,现已逐渐发展为内外因相结合的解释。
综合学派综合生物学派和气候学派的观点,以生物因子和非生物因子间复杂的组合作为种群波动机制的多因性,并因时间、地点的变化而变化。20世纪50年代气候学派与生物学派发生激烈论战,但也有学者提出折中的观点。如美国生态学家米琳(Miline)既承认密度制约因子对种群调节的决定作用,也承认非密度制约因子具有决定作用。他把种群数量动态分成极高数量、普通数量和极低数量。在对物种最有利的典型环境中,种群数量最高,密度制约因子决定种群数量;在环境条件极为恶劣的条件下,非密度制约因子也可以左右种群数量变动。
种群对环境变化的生态对策
编辑在长期的协同进化过程中,生物为了适应其生存的环境逐渐形成了与环境相适应的生态对策。生态对策(bionomic strategies)就是一个物种或一个种群在生存斗争中对环境条件所采取的适应行为,在长期稳定的环境中生活的种群尽可能均匀地利用环境,在迅速出现随后又消逝的环境中,生物能及时地寻找有利的继续生存的地点。1976年,美国生物学家麦克阿瑟(MacArthur)和威尔逊(Wilson)根据生物种群的进化环境和生态对策,可将生物划分为两种对策类型,即r–对策和K-对策。r–对策和K-对策生物的主要特征比较见下表。
r–对策和K-对策生物的主要特征比较
特征 | r–对策 | K-对策 |
适应气候 | 多变,难以预测,不确定 | 稳定,可预测,较确定 |
死亡 | 常是灾难性的、无规律、非密度制约 | 比较有规律,受密度制约稳定 |
种群大小 | 变动大,不稳定,常低于饱和密度 | 稳定,密度临近饱和密度 |
存活 | 存活曲线C型,幼体存活率低 | 存活曲线A、B型,幼体存活率高 |
竞争 | 多变,通常不紧张 | 经常保持紧张 |
选择倾向 | 发育快,增长力强,提早生育,体型小,单次生殖 | 发育缓慢,竞争力强,延迟生育,体型大,多次生殖 |
寿命 | 短,通常小于1年 | 长,通常大于1年 |
最终结果 | 高繁殖力 | 高存活力 |
r–对策者适应于难以预测的多变环境,如干旱地区和寒带。 其是新生境的开拓者,具有能使种群最大化的生物学特征,即高生育力、快速发育、早熟、成年个体小、寿命短等。r–对策者一般具有很强的扩散能力,一有机会就会入侵新的生境并迅速增殖,但由于其把大部分能量用于繁殖,所以一般缺乏保护后代机制,竞争力弱。
K-对策者适应于热带雨林等气候稳定,很少有难以预测的环境。K-对策通常出生率低、寿命长、成年个体大,具有较完善的保护后代机制,一般扩散能力较弱,但由于种群密度较高,竞争较为剧烈。因此,K-对策以“质”取胜,r–对策以“量”取胜。在同一分类单元中,同样可作生态对策比较,如哺乳动物中的啮齿类大部分是r–对策者,而大象、老虎、熊猫等则是K-对策者。
r–对策和K-对策在进化过程中各有其优缺点。K-对策者的种群数量较稳定,生境退化的可能性较小,具亲代关怀、个体大和竞争力强等优点,这使得它们能在生存竞争中取得胜利,但是一旦受到危害,种群数量就会下降。r–对策者虽然防御力弱、无亲代关怀、死亡率较高,但种群可迅速恢复,并离开恶化的原生境,在其他地方建立起新的种群。
在实际环境中,种群对环境的适应不仅有r–对策和K-对策两种方式,还有其他适应方式。如r-K连续选择,即个体越大,世代时间越长,繁殖力越低;反之,个体越小,世代时间越短,繁殖能力越强。此外,还有胁迫忍耐对策,如盐渍环境中的植物就以耐盐种群为主,干旱条件下的植物会通过形态变化来减少水分损失等,以上均是种群应对环境变化采取的生态对策。
种间关系
编辑种间关系(interspecific interaction)是指不同物种种群之间的相互作用。两个种群可以彼此相互影响,也可以互不干扰。如果彼此相互影响的话,这种影响可以是有利的,也可以是有害的,具体关系如下:
两个种群存在的基本相互关系
关系类型 | 物种 | 相互作用的一般特征 | |
A | B | ||
中性(neutralism) | 0 | 0 | 两个种群彼此互不影响 |
竞争(cormpetition) | - | - | 两个种群彼此互相抑制且均受害 |
偏害(amensalism) | - | 0 | 对种群A有害,对种群B无利也无害 |
捕食(predlation) | + | - | 种群A杀死或吃掉种群B中一些个体 |
寄生(parasitism) | + | - | 种群A寄生于种群B并有害于后者 |
偏利(cornnensalism) | + | 0 | 对种群A有利,对种群B无利也无害 |
互利(nutualism) | + | + | 两个种群彼此互相有利 |
在上表中,加号(+)表示有利,减号(-)表示有害,0表示无利也无害。如两个种群互不影响,就可以表达为(0,0),互相有利为(+,+),互相有害为(-,-),对一方有利而对另一方有害则为(+,-),其他的相互关系还有(-,0)、(+,0)等。
种群间的相互关系可分为两类,即正相互作用(positive interaction)与负相互作用(negative interaction)。前者包括偏利作用、互利共生和原始合作,后者包括竞争、捕食、寄生和偏害作用。在生态系统的发育与进化中,正相互作用趋向于促进或增加,从面加强两个作用种群的存活,而负相互作用趋向于抑制或减少。
正相互作用
互利共生
互利共生(mutualism)是指两个物种长期共同生活在一起,彼此相互依赖,相互依存,并能直接进行物质交流的一种相互关系。常见于需求极不相同的生物之间,如豆科(Fabaceae)作物和根瘤菌共生形成的根瘤,其能增加植物从土壤中吸收矿质元素的能力,同时植物也为真菌提供了光合作用的某些产物。另外,动物与微生物间互利共生的例子也很多,如牛的瘤胃内具有密度很高的细菌和原生动物,瘤胃可为其提供生存场所,而这些细菌和原生动物能分解纤维素和纤维二糖,合成维生素,对牛也有利。
根瘤
偏利共生
偏利共生(commensalism)指种间相互作用仅对一方有利,对另一方无影响。附生植物与被附生植物之间是一种典型的偏利共生关系,如地衣(Lichenes)、苔藓(Bryophyte)、某些蕨类(Pteridophyta)等附生在树皮上。在热带森林中还有很多高等的附生植物,这些附生植物借助于被附生植物支撑自己,以获得更多的资源(如光、空间),但对被附生种群则无多大影响。在动物中也存在偏利共生关系,如鮣鱼(Echeneis naucrates)会用头顶上的吸盘固着在鲨鱼腹部,但这仅对鮣鱼有利。
附着在树皮上的苔藓
原始协作
原始协作(protocooperation)是指两种群相互作用,双方获利,但协作是松散的、分离后,双方仍能独立生存。如寄居蟹和某些腔肠动物的共生关系,腔肠动物附着在寄居蟹背上,当寄居蟹在海底爬行时,扩大了腔肠动物的觅食范围。同时,腔肠动物的刺细胞又对蟹起着伪装和保护作用。此外,鸵鸟与马也会进行协作,前者视觉敏锐,后者嗅觉出众,对共同防御天敌十分有利。
寄居蟹
负相互作用
竞争
竞争通常包括两种形式,一为直接干涉型,如动物之间的格斗;二为资源利用型,如水稻(Oryza sativa)与稗草(Echinochloa crus-galli)对阳光、水和养分的竞争。竞争者双方都力求抑制对方,其结果使双方的增长和存活都受到抑制。竞争的结果可向两个方向发展,一是一个物种完全挤掉另一物种,如美国西部的冰草(Agropyron cristatum)被欧洲的雀麦(Bromus japonicus)淘汰;二是不同物种占有不同的空间,捕食不同食物,或其他生态习性上的分离,即生态分离,也可能使两种间形成平衡而生存,如非洲草原狮(Panthera leo)、豹(Panthera pardus)、狼(Canis lupus)等捕食的主要猎物上的分离、森林中不同高度植物种的分离等。
水稻与稗草的竞争阳光
捕食
捕食是指某种生物通过消耗其他生物活体的全部或部分身体,直接获得营养以维持自身生命的现象。前者称为捕食者(predator),后者称为猎物(prey)。捕食是一个种群对另一个种群的生长与存活产生负效应的相互作用,从广义的概念看,捕食还是高一营养级的生物取食和伤害低一营养级的生物的种间关系。
广义的捕食包括传统捕食、同类相食(cannibalism)、植食(herbivory)和昆虫拟寄生者(parasitoid)。传统捕食指食肉动物捕食食草动物或其他食肉动物;同类相食是捕食的一种特殊形式,即捕食者和猎物均属同一物种;植食指动物取食绿色植物营养体、种子和果实,这种情况下植物往往未被杀死而受损害;昆虫拟寄生者是指昆虫界的寄生现象,寄生昆虫常常把卵产在其他昆虫体内,待卵孵化为幼虫以后便以寄主的组织为食以获得营养,直到寄主死亡为止,它们与真寄生虫者的区别是总要杀死其寄主。狭义的捕食仅包括动物吃动物这一情况。
食肉动物捕食食草动物
寄生
寄生是指一个物种(寄生者)寄居于另一个物种(寄主)的体内或体表,从而摄取寄主养分以维持自身生活的现象。根据寄居在寄主身体位置的不同,寄生可以分为体外寄生和体内寄生两类。由于寄生具有一定专一性,因此寄生者和寄主常常是协同进化的,寄生物为适应它们的寄主表现出极大的多样性。寄主可以是植物、动物,也可以是其他寄生物。在植物界,寄生性种子植物还可分为全寄生与半寄生两类。全寄生植物从寄主那里摄取全部营养,而半寄生植物仅仅是从寄主那里摄取无机盐类,它自身能进行光合作用并制造养分。全寄生的高等植物主要有大花草(Rafflesia arnoldii)、菟丝子(Cuscuta Chinensis)等,半寄生的植物有小米草(Euphrasia pectinata)、榭寄生(Viscum coloratum)等。
菟丝子
偏害作用
在自然界中,偏害作用很常见,其主要特征是当两个物种在一起时,由于一个物种的存在,可以对另一物种产生抑制作用,而该物种自身不受影响。异种抑制作用(又称为他感作用)和抗生素作用都属于偏害作用的类型。如胡桃树会分泌一种被称为胡桃醌的物质,它能抑制其他植物的正常生长,从而产生有害的结果。抗生素作用是一种微生物产生一种化学物质来抑制另一种微生物的过程,如青霉素就是青霉菌所产生的一种细菌抑制剂,也就是通常所说的抗生素。
胡桃树分泌胡桃醌
种内关系
编辑存在于生物种群内部个体间的相互关系称为种内关系(intraspecific relationship),生物的种内关系包括密度效应、动植物性行为(植物的性别系统和动物的婚配制度)、领域行为、社会等级、通信行为以及利他行为等。
密度效应
动物
在种内关系方面,动物种群和植物种群的表现有很大区别。许多实验揭示了动物密度效应的各种机制。如当把一对果蝇(Drosophilidae)置于一个瓶中,并供应一定量的食物时,起初其后代数量会迅速增加,很快就会达到一定限度。但当把很多果蝇置于同样的瓶中时,每对果蝇所产出的后代数量将与瓶中果蝇的密度成反比。这种效应是由于幼虫竞争食物引起的,密度越大竞争所引起的死亡率越高,成虫的寿命也会下降,引起成虫寿命下降的密度比引起幼虫存活率下降的密度高。密度制约因素对未成年期的有害影响往往比成年期大。
美洲斑潜蝇(Liriomyza sativae)实验种群密度效应研究表明,卵期不存在密度效应,叶片上的卵量及分布与潜蝇成虫数量的多少有关;幼虫期存在密度效应,1龄幼虫对幼虫期存活率、预蛹重、成虫寿命、繁殖等有显著的影响。密度超过0.8头/立方厘米时,由于空间不足、营养不足、排泄物及脱皮物增多,生态环境恶化,个体发育不良。同时,营养不足又使幼虫间自残(cannibalism)。
植物
植物个体间的密度效应(density effect)反映在个体产量和死亡率上。在一定时间内,当种群的个体数目增加时,就必定会出现邻接个体之间的相互影响,称密度效应或邻接效应(the effect of neighbours)。因为植物不像动物那样逃避密集和环境不良的情况,其表现只是良好情况下可能枝繁叶茂,而高密度下可能枝叶少,构件数少。植物的密度效应有两个特殊的规律,具体如下:
最后产量衡值法则
最后产量衡值法则(Law of constant final yield)指的是不管初始播种密度如何,在一定范围内,当条件相同时,植物的最后产量差不多总是一样的。澳大利亚生态学家唐纳德(Donald)对三叶草(Trifolium subterraneum)密度与产量的关系作了一系列研究后,证实了这个法则。
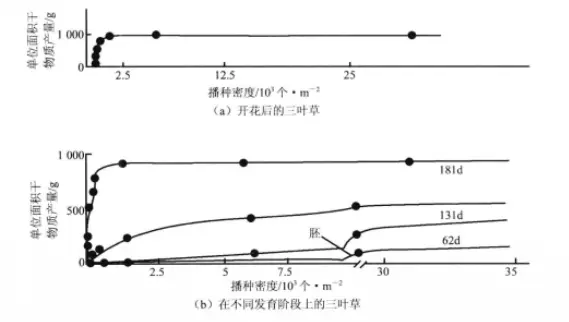

三叶草单位面积干物质产量与播种密度之间关系
上图表示是单位面积上三叶草的干物质产量与播种密度的关系。其中由图(a)可以看出,在密度很低时干物质产量随播种密度增加,但很快就趋于稳定;由图(b)可以看出,从萌芽初期到181天,都呈现出产量随密度恒定的规律。最后产量衡值法的公式为Y(单位面积产量)=W(植物个体平均重量)×d(密度)=Ki(常数),最后产量衡值法则的生物学意义为在稀疏种群中的每一个个体,都很容易获得资源和空间,生长状况良好,构件数多,生物量大;而在密度高的种群中,由于叶子相互重叠,根系在土壤中交错,植株彼此之间对光、水、营养物质等竞争激烈,在有限的资源中,个体生长速度降低,个体变小。
-3/2自疏法则
生长于较高密度种群内的植物,由于密度的抑制作用,种群内个体会逐渐死亡,种群数目逐渐减少,直至达到平衡。这种种群的生长动态现象被称为自然稀疏或者自疏(self-thinning)。自疏现象普遍存在于自然和人工植物种群中,与农业、林业以及牧草生产有着密切的联系。1963年,日本生态学家尤达(Yoda)研究了植物自然种群和栽培作物种群的自疏过程,提出了-3/2自疏法则,表达式为W(种群内植株的平均生物量)=K(由特定物种决定的常数)d(植物种群的密度)。
由-3/2自疏法则可知,在一个空间拥挤、年龄均等、构成单一的植物种群中,随着个体植株生物量的增长,其种群密度将相应减少,且在双对数坐标系里,lgW-lgd的点运动轨迹不会超越一条斜率为-3/2的直线上限。而与植物的生活型、年龄大小、土壤状况及其他栖息地条件等因素无关。-3/2方自疏法则描述了植物种群在自疏过程中,存活个体的平均质量增加与逐渐减小的密度之间动态关系的经验规律,此规律为植物种群所特有,广泛适用于大小悬殊、形态各异的各种植物。
-3/2自疏法则
动植物性行为
植物的性别系统
大多数植物个体是具有雌雄蕊的两性花,有一些植物个体具有雌雄两类花,雄花产生花粉,雌花产生胚珠,所以是雌雄同株而异花的,如玉米(Zea mays)。雌雄异株植物的雌花和雄花分别长在不同的植株上,在植物界中,雌雄异株植物较为稀少,大约只占有花植物的5%左右,如银杏(Ginkgo biloba)。多数生物学家认为,雌雄异株能减少同系交配的几率,具有异性杂交的优越性。此外,雌雄异株实际上是回避两性间竞争的对策,增加了两性利用不同资源的能力,也减少了食种子动物的压力。
银杏雄株
银杏雌株
另一个环境压力是由脊椎动物传粉造成的,如藤思兜树属(Freycinetia)的多数植物为雌雄异株,只有含单性花的穗状花序,但偶尔也会出现雌雄同株的植株,其传粉动物是沙蒙狐蝠(Pteropus samoensis),其在取食有甜味的肉质苞片时,对雄花和两性花的危害比雌花大。当狐蝠在雌雄异体植株上来取食时,雄花序虽然被破坏了,但花粉粘着在狐蝠面部,再转到雌花序采食就使后者授粉,同时对雌花序危害又不大。相反,当狐蝠在雌雄同株的两性花序上来进食时,使其遭到较大的破坏。所以,雌雄同株个体在进化选择上处于劣势,而雌雄异株个体将成为适者而生存下来,藤露兜树沿雌雄异株方向进化。藤露兜树的例子说明了植物性别系统的进化选择中环境因素影响的复杂性和研究的困难性。与高等动物相比,植物性别的特点是其多样性和易变性。同一属或科的植物种中,有的自花授粉,有的异型杂交,而进化中已形成的、防止自花授粉的方式也很多。
藤露兜树
动物的婚配制度
动物的婚配制度是指种群内婚配的各种类型。婚配包括异性间相互识别、配偶的数目,配偶持续时间以及对后代的抚育等。动物的婚配制度从单配到多配,中间还有许多过渡类型。甚至同种动物在不同年份、不同季节、不同地理位置,其婚配制度可能会有变化。因此,一个物种的婚配制度是以一个个体在一个繁殖季节里和其交配的配偶数来定义的。按配偶数划分,婚配制度有单配偶制(monogamy)和多配偶制(polygamy)。
单配偶制是雌雄两性个体彼此独占,这种独占或是直接的,或是通过控制资源而实现的。在动物界内,单配偶制是比较少见的,只有鸟类以一雄一雌制较普遍,如鸳鸯(Aix galericulata)、天鹅(Cygnus)、丹顶鹤(Grus japonensis)和许多鸣禽。哺乳类仅有少数动物为单配偶制,如狐(Vulpes)、河狸(Castor fiber)等。单配偶制主要出现在那些雌雄双亲只有共同养育幼体,才能使繁殖变得有成效的种类,倘若配偶中有一方把能量消耗在与其它个体的婚配上,那就会使幼体失去生存的机会。单配偶制也出现在资源分布均匀,使每一个个体很少有可能独占资源的情况下,此时只有双亲共同担负抚育后代的职责,才能在进化上获得最佳效果。
丹顶鹤
多配偶制是指个体拥有两个或多个配偶,而这些配偶中没有与其它个体再成配偶的,多配偶制具体可分为以下3种类型:
一雄多雌制(polygyny)是指一个雄性占有多个雌性,是最常见的婚配制度,又可分为四类:保卫资源型,如蜂鸟(Trochilidae);保卫雌性型,如海豹、野马;雄性优势型,如黑琴鸡(Lyrurus tetrix);高速多窝型为雄鸟参与孵化和育雏的类型,如一些鸡类。
蜂鸟
一雌多雄制(polyandry)是指一个雌性占有多个雄性,是较为稀见的一种动物婚配制度。其前提是雄性能担负孵化和育雏的职责,使雌性能多产几窝。另一个条件是雌性有进行多次交配的能力,并能抚育幼体。一雌多雄制也有两种类型,一类为保卫资源型,如距翅水雉(Jacana spinosa),其雌鸟常与多只雄鸟交配,在不同地方产卵,因此可见到其领域内同时有多只雄鸟伏窝的现象,雌鸟对卵和雏鸟则很少照料。另一类为雌性控制型,如红颈瓣蹼鹬(Phalaropus lobatus),其雌鸟体色比雄鸟鲜艳,会为了雄鸟进行竞争。
距翅水雉
混交制是指无论雌雄都可以与一个或更多的异性交配,而不形成相对固定的婚配关系,如鸟类中约有6%的种类是混配制的,哺乳动物如生活在苏格兰东北部的里氏田鼠(Microtus richardsoni)。
里氏田鼠
领域行为
动物个体、配偶或家族通常都只是局限活动在一定范围的区域。如果这个区域受保卫,不允许其他动物,通常是不让同种动物的进入,那么这个区域或空间就称为领域(territory),而动物占有领域的行为则称为领域行为或领域性(territoriality)。领域行为是种内竞争资源的方式之一,占有者通过占有一定的空间而拥有所需要的各种资源。领域行为常见于大部分脊椎动物和一些节肢动物种类,这与它们具有筑巢和育幼的行为特性有关。在脊椎动物当中,鸟类的领域行为最为发达,啮齿类动物和猿猴类的群体领域最为常见。节肢动物中蜻蜓(Aeshna melanictera Selys)、蟋蜂、各种膜翅目昆虫、蝇类等也有领域行为。
根据领域性质的不同,领域可以划分为繁殖领域(所保卫的区域用于求偶、交配、筑巢和繁殖)和非繁殖领域(如觅食领域)。领域的特征主要有以下三个方面:1. 排他性。领域由一个个体或几个个体(一对配偶、一个家庭或一个成员相互之间没有亲缘关系的群体)独占使用,不允许其他动物,通常是同种动物的进入,领域占有者会积极保卫自己的领域;2. 伸缩性。领域范围的大小取决于物种。在同一物种内,领域的大小随着生态条件或时间的变化而有所调整;3. 替代性。当领域的占有者被移去或自然死亡后,它们的领域很快地就被其他新来者所占领。
社会等级
社会等级(social hierarchy)是指动物种群中各个动物的地位具有一定顺序的等级现象,社会等级形成的基础是支配行为,或称支配-从属(dorminant-submissive)关系。如家鸡,开始时鸡群中存在彼此啄击、打斗、威吓等现象。经过一段时间后,就会逐渐形成等级,稳定下来后,低级的一般表示妥协和顺从,但有时也通过再次格斗而改变顺序等级。稳定的鸡群往往生长快,产蛋也多,其原因是不稳定鸡群中个体间经常的相互格斗要消耗许多能量。
家鸡
社会等级制在动物界中相当普遍,包括许多鱼类、爬行类、鸟类和兽类。高地位的个体通常较低地位的从属个体身体强壮、体重大、性成熟程度高,具有打斗经验,其生理基础是血液中有较高浓度的雄性激素。一般说来,社群中雌雄各有等级顺序,主雄多与主雌或若干强雌交配,不允许其他雄体与后者交配。
社群等级和领域性这两类重要的社会性行为,与种群调节有密切联系。英国生态学家温·爱德华兹(Wyhne·Edwards)的种群行为调节学说基础就是这种社会性行为与种群数量的关系,当动物数量上升很高时,全部最适的栖息地被优势个体占满。次适地段虽然能成为其余个体的适宜栖息地,或者说可栖息密度具有一定弹性,但这种弹性有一定限度。随着密度增高,没有领域或没有配偶的从属个体比例也将增加,它们最易受不良天气和天敌的危害,这部分(称为剩余部分)比例的增加意味着种群死亡率上升,出生率下降,限制了种群的增长。相反,当种群密度下降时,这部分比例上升,种群死亡率降低,出生率上升,促进了种群的增长。
通信行为
通信(communication)指的是个体通过释放一种或是几种刺激性信号,引起接受个体产生行为反应。根据信号的性质和接受的感官,可以把通信分为视觉、化学和听觉等。信息传递的目的很广,如个体识别(包括识别同种个体、同社群个体、同家族个体等)亲代和幼仔之间的通信,两性之间求偶,个体间表示威吓、顺从和妥协,相互警报,标记领域等。从进化意义来说,所选择的通信方式应以传递方便、节省能耗、误差小、信号发送者风险小及对生存必需的信号。
利他行为
利他行为(altruism)指的是个体牺牲自我而使社群整体或其他个体获得利益的行为,例如,当白蚁的巢穴被打开时,工蚁和幼虫都向内移动,兵蚁则向外移动以围堵缺口,表现出保卫群体的利他行为。工蜂在保卫蜂巢时放出毒刺,这实际上是一种“自杀行动”。亲代关怀(parental care)也是一种利他行为,亲代为此要消耗时间和能量,但能提高后代的存活率。一些鸟类当捕食者接近其鸟巢和幼鸟时佯装受伤,以吸引捕食者追击自己而将其引开鸟巢,然后自己再逃脱。不少啮齿类在天敌逼近时发出特有鸣叫声或以双足敲地,向周围的鼠发出报警信号,而发信号者反而更易引起捕食者的注意。
工蜂保卫蜂巢
参考资料
编辑展开[1]闫云君主编, 徐莉, 唐朝晖副主编. 生命科学导论 医学版 第3版[M]. 2020: 151-162. (12)
[2]张闻, 郑多编. 全国普通高等医学院校五年制临床医学专业十四五规划教材 医学生物学 第2版[M]. 北京: 中国医药科学技术出版社, 2022: 203-204. (2)
[3]赵玉山, 高丽芳主编, 《科学新探索读本》丛书编写组编写. 生物群落的探究[M]. 2011: 7-9. (3)
[4](英)乔治B. 生物学的思维方式 第9版[M]. 北京: 北京联合出版公司, 2022: 787. (2)
[5]张坤, 张颖, 李永峰主编, 张宝玺副主编, 程国玲, 李伟军主审. 基础生态学[M]. 哈尔滨: 哈尔滨工业大学出版社, 2018: 41. (4)
[6]梁士楚, 李铭红主编, 聂呈荣, 王文强, 饶本强等副主编. 生态学[M]. 武汉: 华中科技大学出版社, 2015: 76-83. (8)
[7]程胜高, 罗泽娇等主编. 环境生态学[M]. 北京: 化学工业出版社, 2003: 28.
[8]符裕红主编. 生态学基础实验教程[M]. 北京: 中国农业大学出版社, 2020: 67.
[9]沈显生编著. 生态学简明教程[M]. 合肥: 中国科学技术大学出版社, 2012: 83. (2)
[10]胡荣桂主编. 环境生态学[M]. 武汉: 华中科技大学出版社, 2010: 40-41,50. (6)
[11]骆世明主编. 普通生态学[M]. 北京: 中国农业出版社, 2005: 75-76. (2)
[12]胡志东, 任继文主编. 环境生态学 上[M]. 沈阳: 白山出版社, 2003: 74-75. (2)
[13]《国家教师资格考试专用系列教材》编委会编著. 生物学科知识与教学能力 高级中学 2014-2015[M]. 2014: 91-94. (4)
[14]吴国凯, 秦德智, 古立秀. 生态遗传学[M]. 北京: 读物出版社, 1992: 55.
[15]孙丽主编. 生态学基础 2006年最新版[M]. 天津: 南开大学出版社, 2006: 30-31. (2)
[16]顾卫兵主编. 环境生态学[M]. 北京: 中国环境出版社, 2014: 39. (2)
[17]任南琪等编著. 污染控制微生物学[M]. 哈尔滨: 哈尔滨工业大学出版社, 2002: 33-34. (5)
[18]蔡立哲编. 滨海湿地环境生态学[M]. 厦门: 厦门大学出版社, 2020: 87-88. (3)
[19]邓小华主编. 环境生态学[M]. 北京: 中国农业出版社, 2006: 56-57. (2)
[20]田清涞编著. 普通生物学[M]. 北京: 海洋出版社, 2000: 511-512. (3)
[21]杨玉红, 王锋尖主编, 曾镭, 王剑副主编, 王学英, 孙秀青, 王增池等参编. 普通生物学[M]. 武汉: 华中科技大学出版社, 2012: 170. (2)
[22]陈阜主编. 农业生态学[M]. 北京: 中国农业大学出版社, 2002: 50. (2)
[23]张海林主编. 农业生态基础[M]. 北京: 中央广播电视大学出版社, 2008: 30.
[24]张坤, 张颖, 李永峰主编, 张宝玺副主编, 程国玲, 李伟军主审. 基础生态学[M]. 哈尔滨: 哈尔滨工业大学出版社, 2018: 57-58. (3)
[25]孔繁德主编, 冯雨峰, 刘传才副主编. 生态学基础 第2版[M]. 北京: 中国环境科学出版社, 2011: 75-82. (8)
[26]李维炯主编, 北京师范大学成人高考教材组编. 生态学基础[M]. 北京: 北京邮电大学出版社, 2005: 83-84. (6)
[27]胡志东, 任继文主编. 环境生态学 上[M]. 沈阳: 白山出版社, 2003: 96-97. (3)
内容由百科小编提供,本内容不代表globalbaike.com立场,内容投诉举报请联系全球百科客服。如若转载,请注明出处:http://globalbaike.com/9532/