啮齿目(Rodentia),隶属于脊索动物门哺乳纲,是哺乳纲下物种种类最丰富、数量最多的目之一。根据ITIS(2015年)标准,将啮齿目下分5亚目34科512属2367种。本目动物的上下颌均只有1对门齿,上门齿肥大凿形,虽到成年门齿也无齿根,终生生长,故需不时咬啃磨蚀。啮齿目动物分布在除南极洲、新西兰和部分海岛以外的世界各地,可生活于草原、森林、农田、荒漠、沼泽等多种生态环境中,也可以在人类社区中生活。
起源演化
编辑起源
啮齿目动物的牙齿结构是辨认这类哺乳动物化石的主要特征,其最早的化石记录出现在古新生代。在劳亚古陆(注:板块学说中包括亚洲、欧洲、北美洲的古大陆)多处被发现的化石证据表明,啮齿目动物起源于约6500万年前的白垩纪-第三纪大规模灭绝之前,最早可追溯到7400万年前。然而根据分子生物学分子钟的推算,啮齿目动物最早可能在大约1.1亿年前的晚白垩世晚期出现,这一结果与已知的化石记录相差甚远。


各种啮齿类的上臼齿结构
中国发现的古新生代化石表明,啮齿目动物的起源可能与亚洲特有的宽臼兽类(eurymylids)如晓鼠(Heomys)有关。然而,对于宽臼兽类这一大类的分类和定位仍存在争议,许多学者认为晓鼠和其他一些宽臼兽类不是真正的啮齿目动物。中国古新生代地层中发现的钟健鼠(Cocomys)和外鼠(Exmus),尽管它们的出现时代远晚于北美的副鼠类化石记录,但在系统发育上却更接近整个啮齿目动物的基干,这表明啮齿目动物的起源可能在亚洲,只是尚未发现更早期的化石。
因此,关于啮齿目动物起源的问题在学术界仍存在争议。有观点认为啮齿目动物可能起源于灵长目(Primates)的近兔猴类,也有观点基于跟骨结构怀疑它们可能源自古肉食类动物。
分化与扩张
在6600万年前的白垩纪-古新世灭绝事件中,亚洲多瘤齿兽目(Multituberculata)的种群受到了严重影响,与它们在北美洲和欧洲的近亲不同,该种群一直未能完全恢复。在由此产生的生态空白中,啮齿目动物和其他胎盘哺乳动物得以进化和多样化,填补了多瘤齿兽目留下的生态位。
在白垩纪-古新世边界之后的几百万年内,啮齿目动物与其他胎盘哺乳动物分化开来。随后,在新生代时期,啮齿目动物迅速进行辐射进化。其多样化在整个始新世时期(约5500万年前至3400万年前)都有化石发现,到了大约2000万年前,许多现代可识别的啮齿目动物下的科级分类,如鼠科,已经出现。
啮齿目动物在各大洲的扩张历史非常复杂。副鼠科(Paramyidae)大约在晚古新世/早始新世界限(约5500万年前)扩散至欧洲和亚洲。海狸(Castor fiber)最早出现在晚始新世的欧亚大陆,然后在晚中新世扩散到北美洲。始新世晚期,起源于亚洲的豪猪亚目(Hystricomorpha)动物侵入非洲。
根据非洲的化石证据,豪猪亚目中的豚鼠小目(Caviomorpha)动物又借助海洋洋流漂浮的碎屑横越大西洋,散布到当时孤立的南美洲。在渐新世早期,豚鼠小目抵达了大安的列斯群岛,这表明它们在南美洲实现了迅速扩散。而鼠总科(包括仓鼠科 Circetidae、鼠科 Muridae)的扩散可能涉及多次进化,其中包括在非洲的七次扩张、在北美洲的五次扩张、在东南亚的四次扩张、在南美洲的两次扩张以及在欧亚大陆多达十次的扩张。
命名与分类
编辑命名
啮齿目的学名“Rodentia”,是英国旅行家和自然学家托马斯·爱德华·鲍迪奇(Thomas Edward Bowdich)于1821年首次提出的。“Rodentia”这个现代拉丁词汇来源于rodens,是rodere的现在分词,意为“啃咬”、“咬食”。
分类
啮齿目动物的分类主要根据咬肌和牙齿形态(包括门齿釉质层构造),但意见极为分歧。最初分单门齿亚目(Simplicidentata)和双门齿亚目(Duplicidantata)两个亚目,前者即啮齿目,后者为兔形目。兔形目划出后,啮齿目内的分类更显复杂。啮齿目是哺乳动物中种类最多、数量最大的类群之一。啮齿目分类简单时仅分两个亚目,如松鼠型下颌亚目和豪猪型下颌亚目,或五脊齿亚目和非五脊齿亚目。复杂者可多达16个亚目及许多科或超科。但传统的分法多分为松鼠亚目、鼠形亚目及豪猪亚目3个亚目,或加一始啮亚目成4个亚目。根据ITIS(2015年)标准,将啮齿目下分5亚目34科512属2367种,详细如下表:
亚目 | 科 | ||
中文名 | 学名 | 中文名 | 学名 |
鳞尾松鼠亚目 | Anomaluromorpha | 鳞尾松鼠科 | Anomaluridae |
跳兔科 | Pedetidae | ||
河狸亚目 | Castorimorpha | 河狸科 | Castoridae |
衣囊鼠科 | Geomyidae | ||
异鼠科 | Heteromyidae | ||
豪猪亚目 | Hystricomorpha | 栉趾鼠科 | Ctenodactylidae |
华毛鼠科 | Abrocomidae | ||
滨鼠科 | Bathyergidae | ||
硬毛鼠科 | Capromyidae | ||
豚鼠科 | Caviidae | ||
毛丝鼠科 | Chinchillidae | ||
栉鼠科 | Ctenomyidae | ||
兔豚鼠科 | Cuniculidae | ||
刺豚鼠科 | Dasyproctidae | ||
硅藻鼠科 | Diatomyidae | ||
长尾豚鼠科 | Dinomyidae | ||
针鼠科 | Echimyidae | ||
美洲豪猪科 | Erethizontidae | ||
海地岛鼠科 | Heptaxodontidae | ||
豪猪科 | Hystricidae | ||
河狸鼠科 | Myocastoridae | ||
八齿鼠科 | Octodontidae | ||
岩鼠科 | Petromuridae | ||
藤鼠科 | Thryonomyidae | ||
鼠亚目 | Myomorpha | 跳鼠科 | Dipodidae |
丽仓鼠科 | Calomyscidae | ||
仓鼠科 | Cricetidae | ||
鼠科 | Muridae | ||
马岛鼠科 | Nesomyidae | ||
猪尾鼠科 | Platacanthomyidae | ||
鼢鼠科 | Spalacidae | ||
松鼠亚目 | Sciuromorpha | 山河狸科 | Aplodontiidae |
睡鼠科 | Gliridae | ||
松鼠科 | Sciuridae | ||
形态特征
编辑外形特征
啮齿目动物通常很小(小至10克),但最大的啮齿目动物可达将近70千克(如水豚)。其被毛结构和毛色变化复杂而多样,但各类群的毛色比较稳定,多作种属的鉴别特征。被毛具保护毛(guard hair)和基毛(ground hair)两类。保护毛中以针毛(awns)为主,其毛干粗硬,毛基细而柔软,被毛表面颜色主要决定于针毛;刺毛(bristles)的毛干刚硬,色素较多,散在分布于针毛间,家鼠属(Rattus)多有刺毛,尤以针毛黄鼠(Rattus huang)为多,此外还有非常特化的棘(spines),豪猪科皆有之。基毛中以软毛(fur)为主,毛短而较软,构成很厚的毛层,毛基颜色主要来自软毛和针毛的基段。柔毛(wool)细长而弯曲,十分柔软。
啮齿目动物具乳头2-10对,以4对居多。前肢短,5指(趾)居多,有些退化成4指、3指或1指,指(趾)端具爪。掌和蹠均有肉质垫称之为胼胝。阴茎有阴茎骨,胼胝和阴茎的形态结构均有种属特异性。睾丸腹位、鼠鼷位或进入阴囊。啮齿目动物尾的变化发展与其生活习性密切相关联,变异性大,长短不一,裸露或覆以发达的簇毛;有些类群覆盖排列成环的鳞片,环间丛生刺毛。
不同毛色的水豚
|
|
|
|




解剖学特征
啮齿目主要的分类是依据咬肌和牙齿形态,其中,啮齿类头骨咬肌分为四种基本结构:始啮亚目型、松鼠亚目型、豪猪亚目型和鼠形亚目型。始啮亚目的咬肌限于颧弓,松鼠亚目的咬肌前端深达吻部,豪猪亚目的深部咬肌穿过眶下孔,鼠形亚目是松鼠亚目和豪猪亚目的结合。
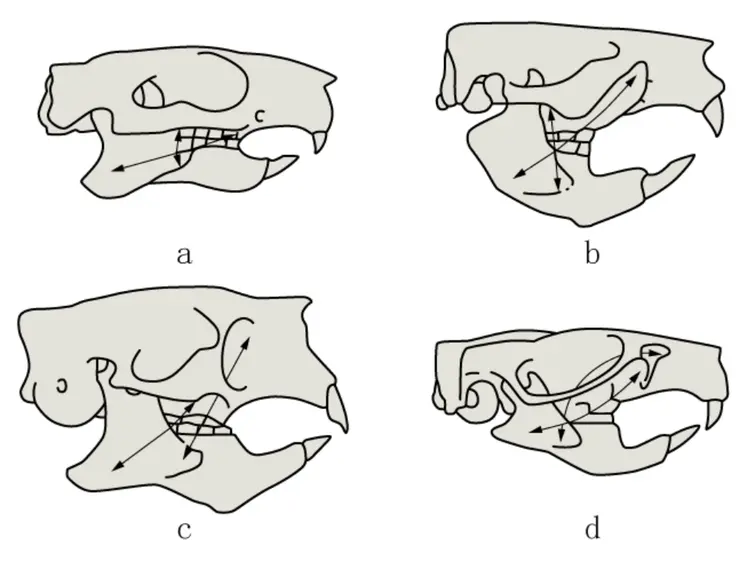
啮齿类头骨咬肌的四种基本结构。a:始啮亚目,b:松鼠亚目,c:豪猪亚目,d:属形亚目
啮齿目动物颅骨的颧面部短于脑颅部,多数仅为脑颅部之半。脑颅平滑或有脊,如额脊、颞脊、矢状脊和人字脊。鼻骨明显。间顶骨存在,但亦有与顶骨或上枕骨愈合。颅两侧咬肌(massete)特别发达,由于与不同咬啮方式适应,致使颧骨和上颌骨有形态结构的不同,颧弓完全向两侧伸展,颧骨多有颧板,供咬肌附着,在原始类群如松鼠的眶下孔窄,其外壁有明显的咬(嚼)肌结节。鼓骨形成听泡,其大小和两者间的距离在不同种属间有显著差异。听泡后缘连乳骨,向腹面伸出有乳突,外枕骨腹侧有颈突或称副乳突,乳突和副乳突的发达程度在各科属间有一定差异。眼眶敞开,并不以骨片联成骨环,眼窝与颞窝相通。眶上突在鼯鼠科和松鼠科显著。腭骨发达,前缘与前颌骨接壤处有门齿孔,是血管和神经的通道。
本目动物上下颌均只有1对门齿,上门齿肥大凿形,虽到成年门齿也无齿根,终生生长,故需不时咬啃磨蚀。无犬齿,留下很大的虚位称齿隙,前臼齿不超过2/1,臼齿3/3、总齿数一般不超过22枚。咀嚼面的珐琅质伸入齿质构成坚硬的齿面(冠),恒定而复杂的齿尖(突)和齿脊并呈现特异图形,为分类鉴定的重要依据。如鼠科的成3纵列丘状齿突(3叶状珐琅质环)或左右对称的2纵列丘状齿突;有些种类又形成复杂齿脊。上颌和下颌左右齿列间的距离相对应,咀嚼时下颌会向前后或斜向移动。
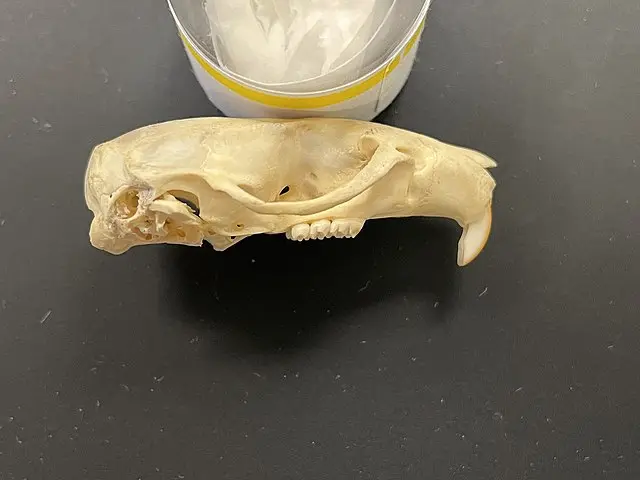
啮齿动物头骨
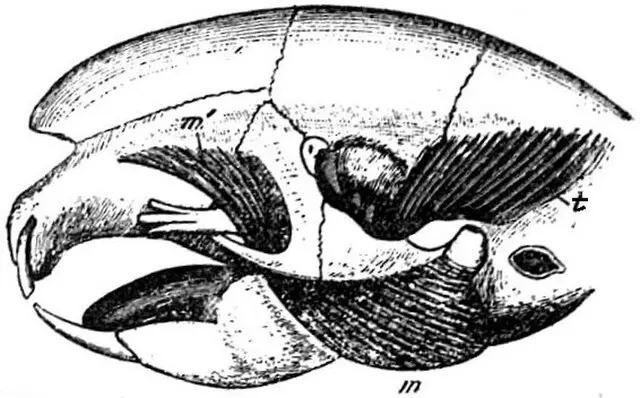
豪猪(Hystrix cristata)头骨

水豚臼齿
分布栖息
编辑分布范围
啮齿目动物分布于除南极洲、新西兰和一些海洋岛屿以外的世界各地。
栖息环境
在进化过程中,啮齿目动物能广泛地适应不同生活条件,一般说来栖息地没有严格的选择性。分布也较广泛,能够栖息于草原、森林、农田、高山、丘陵、平原、荒漠、沼泽、居民区等各种环境中。生态类型多样,有地栖、树栖、半树栖半地栖、地下生活、半地下半地面生活、水栖、半水半陆栖等类型。至于家栖性啮齿目动物,可以说凡是人类所到的地方,就有它们的存在。该目动物几乎可以生存于中国现有的任何一种类型的栖息地中,从喜马拉雅山脉和青藏高原的高海拔地区到热带雨林、农田、城市,甚至停泊在港口的船舶上都有啮齿目动物的存在。

北松鼠栖息于树上

雪地中栖息的东美松鼠(Sciurus carolinensis)

森林中的蓝腹松鼠(Callosciurus pygerythrus)
生活习性
编辑活动特点
啮齿目动物性怯弱,行动敏捷,有些种类营日间活动,有些种类只在夜间活动,夜间活动的鼠类,在某些特殊情况下(如极度饥饿、或受惊扰)偶尔在日间活动。一般鼠类的活动,只限于其洞穴的附近几里地以内,不作长距离的移动,但由于某种原因(人类活动、食物缺乏)亦常使鼠类迁移其洞穴。例如黄鼠(Spermophilus)在盛夏草高时,则喜迁移到耕地中活动。家栖性鼠类,通常会跟随人及交通工具旅行各地。花鼠(Tamias sibiricus)的活动集中在早晨,喜欢在地面活动,但也爬树。
适应能力
啮齿目对环境亦有各种各样的适应能力。如栖息于大陆性气候地带的咽齿类,每年要进行更换体毛1~3次,以适应各种季节不同气候,它们所营的洞穴,一方面可以避寒,另一方面也可以避暑。如黄鼠则可以夏蛰冬眠的形式来渡过不利于生活的季节。
觅食行为
大多数咽齿目动物是典型的草食动物,以植物的茎、根、果实为主要食料。但在有条件时它也食肉类,甚至昆虫。尽管啮齿目动物传统上被认为是食草动物,但大多数小型啮齿目动物都会机会主义地在其饮食中加入昆虫、蠕虫、真菌、鱼或肉,而其中一些已经专门依赖于动物性食物。功能形态学研究啮齿动物的牙齿系统支持了原始啮齿动物更多是杂食性而不是纯粹的食草动物的观点。
例如褐家鼠(Rattus norvegicus)比较偏于肉食甚至盗食小鸡、小鸭。黑线姬鼠(Apodemus agrarius)的食性较广,凡是谷物种子、草籽、豆类,甚至对昆虫也不拒食。倭松鼠(Tamiops maritimus)主要以各种种子、嫩芽、板栗、地衣、树皮和昆虫为食。有时亦食植物的花蜜。来自北美干旱地区的食蝗鼠(Onychomys torridus)以昆虫、蝎子和其他小老鼠为食,仅极少部分食用植物材料。它具有粗胖的身体、短腿和尾巴,但非常灵活,可以轻松地捕食与自身大小相当的猎物。

正在觅食的黑线姬鼠

在树间觅食的倭松鼠
冬眠行为
啮齿目动物主要在隧道(洞穴)中冬眠,隧道的特点是具有适宜其生活的气候。如花鼠9月至翌年3-4月冬眠,雌性率先进入冬眠,雄性随后。在冬季,啮齿目动物会休息并且以其储存的食物为食,或者在雪下活动。一层只有几厘米高的雪就可以保护一些啮齿目动物在至少零下6°C下不会被冻死,而30厘米的雪可以保护它们不会受到严重的或者长时间的霜冻,并且有助于繁殖。但不是所有的啮齿目动物都冬眠,如北松鼠和岩松鼠(Sciurotamias davidianus)无冬眠习性。

冬眠的榛睡鼠(Muscardinus avellanarius)
社会行为
啮齿目动物展现出多样的社会行为类型。如裸鼹鼠(Heterocephalus glaber)是被发现的第一种真正具有社会性的哺乳动物。它们的社会结构与蚂蚁、白蚁以及一些蜜蜂和黄蜂相似。在裸鼹鼠中,只有1只雌性(女王)和1~3只雄性参与繁殖,而其他群体成员则担任劳动者的角色。
此外,又如高度社会化的草原犬鼠属(Cynomys)动物生活在大型家庭群体中,这些家庭群体是它们社会结构的基本单位。同一家庭群的成员共享相同的领地,它们通过口头接触、亲吻和相互梳理来互动。但它们不会对来自其他家庭群体的草原犬鼠执行这些行为。
相比之下,睡鼠属(Glis)动物则过着独立的生活方式。成年睡鼠可能有重叠的觅食范围,但它们生活在个体巢穴中并分开觅食,只在繁殖季节短暂地汇聚以交配。
交流行为
嗅觉
啮齿目动物的主要交流方式是嗅觉。啮齿目类动物以尿液、粪便和皮脂的形式留下化学信号,这些化学信号来自多种专门的气味腺,以此来传达它们的状态、存在、繁殖、身份、社会地位以及其他特征情况,甚至可以用来被捕食者定位猎物。
啮齿目动物能够通过气味来识别近亲,从而表现出亲属优势行为(对亲属的优先行为)以及避免近亲交配。这种亲属识别是通过尿液、粪便和腺体分泌物中的气味线索实现的。主要的评估可能涉及到主要组织相容性复合物(MHC),两个个体的亲缘程度与它们共有的MHC基因相关。在非亲属交流中,需要更持久的气味标记,比如领地边界,这时可能也会使用不挥发的主要尿液蛋白(MUPs),它们充当信息素传递者的功能。MUPs还可以传递个体身份信息,每只雄性家鼠(Mus musculus)的尿液中含有大约十几种基因编码的MUPs。它们通过排尿来标识领地、个体和群体,以及社会组织。
欧亚红松鼠(Sciurus vulgaris)会调查并熟悉邻居的气味,对其侵入的反应不如对非领地的“漂泊者”或陌生人那么激烈。这被称为“亲敌效应”。
听觉
许多啮齿目动物,尤其是那些白天活动且社会性的物种,当它们感知到威胁时,会发出各种警报叫声。这么做有直接和间接的好处。潜在的捕食者可能会在知道自己已被发现时停下来,或者警报叫声可以让同种群或亲缘关系较近的个体采取规避行动。例如,一些物种,如草原犬鼠属拥有复杂的防捕食者警报叫声系统。这些物种可能针对不同的捕食者(例如空中捕食者或地面捕食者)发出不同的叫声,每个叫声都包含有关特定威胁的性质的信息。叫声的声音属性还传达了威胁的紧急程度。
社会性的啮齿目动物比独居物种具有更广泛的叫声范围。 如 Fukomys micklemi(尚无确凿中文名)成年个体有十五种不同的叫声类型,幼年个体有四种不同的叫声类型。类似地,灌丛八齿鼠(Octodon degus)等其他社会性的穴居啮齿动物展示出丰富的交流方式,其叫声库包括十五种不同声音类别。超声波叫声在榛睡鼠属(Muscardinus)之间的社会交流中扮演着一定的角色,尤其是在个体们彼此看不见的情况下使用。
视觉
与除灵长类动物外的所有胎盘哺乳动物一样,啮齿目动物的视网膜中只有两种类型的光感受锥体,即短波长的“蓝-紫外”类型和中波长的“绿”类型。因此,它们被分类为二色视动物;然而,它们在紫外(UV)光谱范围内具有视觉敏感性,因此可以看到人类无法看到的光。例如,在灌丛八齿鼠中,腹部反射更多的紫外光,而背部反射较少。因此,当灌丛八齿鼠感到惊慌时,会站在后腿上,暴露腹部给其他灌丛八齿鼠,紫外视觉可能在传达惊慌信息方面起到作用。当它四脚着地时,它的背部反射的紫外光较少,有助于使灌丛八齿鼠对捕食者不太可见。
触觉
振动行为在啮齿目动物中发挥重要作用,用于提供有关特定行为、捕食者警告、群体或群组维护以及求偶等方面的信息。中东盲鼹鼠(Nannospalax ehrenbergi)是第一种被记录为使用地震通讯的哺乳动物。这些穴居啮齿类动物会用头撞击隧道壁。最初认为这种行为是它们挖掘隧道的一部分,但后来人们意识到它们通过生成有时间模式的地震信号,与邻近的中东盲鼹鼠进行远距离通讯。
脚踏行为是穴居啮齿目动物和半穴居啮齿目动物中常见的捕食者警告和自卫行为。例如,旗尾更格芦鼠(Dipodomys spectabilis)在各种情境下会发出多种复杂的脚踏鼓声,其中包括遭遇蛇类时。这种脚踏行为可以提醒附近的幼仔,同时也传达出鼠类极度警惕,以防止蛇类的袭击。此外,脚踏行为还与雄性之间的竞争有关,优势雄性通过踏击来显示其资源保持潜力,从而减少与潜在对手的实际接触。
家域行为
当生存空间变得有限时,雌性会杀死群体中具有竞争性的其它雌性的幼仔,以此来争夺洞穴、巢穴、食物和后代生活空间。因此,领地性是雌性啮齿目动物用来保护其幼仔免受伤害的一种策略。北松鼠家域2-10公顷,常有重叠,雌雄两性都在家域中营造一个或多个窝。
认知能力
啮齿目动物具有先进的认知能力。它们可以很快学会避开毒饵,这使得它们成为难以对付的害虫。例如,豚鼠(Cavia porcellus)能够学习和记住通往食物的复杂路径。松鼠属(Sciuridae)和跳鼠科(Dipodidae)动物能够通过空间记忆来找到食物的储藏地,而不仅仅依靠嗅觉。
生长繁殖
编辑交配制度
一雄一雌制
啮齿目动物中,有些物种是一雄一雌制的,成年雄性和雌性形成持久的配对关系。这种一雄一雌制可以分为两种形式:强制性和选择性。
在强制性一雄一雌制中,雌雄双方都照顾后代,并在后代的生存中发挥重要作用。这种情况发生在加州鹿鼠(Peromyscus californicus)、灰背鹿鼠(Peromyscus polionotus)、马岛鼠(Hypogeomys antimena)和海狸等物种中。在这些物种中,雄性通常只与自己的伴侣交配。除了对幼仔提供更多的关怀之外,强制性一雄一雌制对成年雄性也有益处,因为它减少了找不到配偶或与不育雌性交配的机会。
在选择性一雄一雌制中,雄性不提供直接的育儿照顾,与一名雌性保持在一起是因为它们在空间上分散分布,无法接触其他雌性。草原田鼠(Microtus ochrogaster)应该是这种一雄一雌制的例子,雄性会保卫和捍卫附近的雌性。
一雄多雌制
而在一雄多雌制的物种中,雄性会努力垄断并与多个雌性交配。与一雄一雌制相似,啮齿目动物中的一雄多雌制种类也有两种形式:防卫型和非防卫型。
防卫型一雄多雌制包括雄性控制拥有吸引雌性的资源的领地。这种情况发生在如黄腹旱獭(Marmota flaviventris)、加州黄鼠(Otospermophilus beecheyi)、哥伦比亚地鼠(Urocitellus columbianus)和理查森地鼠(Urocitellus richardsonii)中。拥有领地的雄性被称为“定居”雄性,而居住在这些领地内的雌性则被称为“定居”雌性。例如,黄腹旱獭的定居雄性似乎永远不会失去它们的领地,它们总是在与入侵雄性的争斗中获胜。一些物种还以直接保卫领地内的雌性,导致激烈的争斗,可能会导致严重伤害。
在非防卫型一雄多雌制物种中,雄性不占有领地,它们四处游荡以寻找雌性来垄断交配机会。这些雄性建立支配等级,排名高的雄性可以接触更多的雌性。这种情况发生在贝尔丁地鼠(Urocitellus beldingi)和一些树松鼠物种中。
选址筑窝
啮齿目动物的幼仔出生时,根据物种的不同,可能表现为早熟状态(多毛、睁眼和相对发育完全)或晚熟状态(闭眼、无毛和相对发育不全)。松鼠属和家鼠等物种通常属于晚熟状态,而豚鼠和豪猪(Hystrix hodgsoni)等物种则通常是早熟的。
在晚熟状态的物种中,雌性在分娩前通常会建造精致的巢穴,并维持巢穴,直到幼仔断奶。雌性通常是坐着或躺着分娩,幼仔会从母兽面对的方向出现。新生幼仔通常会在睁开眼睛几天后首次离开巢穴,最初会定期返回。随着幼仔的成长和发育,它们会逐渐减少回巢,最终在断奶后永久离开。
而在早熟状态的物种中,母兽很少或甚至根本不会建造巢穴。分娩时,雌性通常站立,幼仔会从母兽身后出现。这些物种的母兽通过母兽联系叫声与高度活跃的幼仔保持联系。尽管这些早熟幼仔相对独立,几天内就会断奶,但它们可能会继续接受母兽的哺乳和梳理。
啮齿目动物平均窝大小通常是可用乳房数量的一半。倭松鼠窝常筑于树上树权或树洞内。大足鼠(Rattus nitidus )的洞穴多在荆棘灌丛和岩石缝隙中。
产仔育幼
啮齿目动物的繁殖能力较强,年繁殖1-2胎,多者可达8-10胎,每胎产2-10余仔。啮齿目动物的窝产仔数与乳腺数量呈显著正相关,最大窝产仔数近似于母兽乳房数量。例如家鼠(Mus musculus)怀胎期只20日左右,每年可繁殖6-7次,每次产幼鼠6-7只,最高可达12只以上,幼鼠成熟只需一个月,母鼠在哺乳期仍可照常交尾受孕。豪猪每年1胎,妊娠期大约110天,每胎产1-3仔。河狸妊娠期为106天左右,每胎1-6仔,哺乳期约2个月,幼仔出生后2天就会游泳,第三年性成熟。复齿鼯鼠(Trogopterus xanthipes)每胎产仔1-4只,妊娠期78-89天,22日左右达到性成熟。
凭借强大的繁殖能力,再加上栖息环境可以持续提供足够的食物和有利的气候因素,时有发生特定地区啮齿动物种群数量过剩,甚至导致自然平衡被破坏。

出生8天的小毛足鼠(Phodopus roborovskii)

正在哺乳的水豚
啮齿目动物母兽既提供直接的亲代照顾,如哺乳、梳理、检索和挤在一起保暖,又提供间接的亲代照顾,如食物储藏、筑巢和保护后代。在许多社会性物种中,幼崽可能由非双亲的个体照顾,这一做法称为异养或合作育雏。这种情况已知发生在黑尾土拨鼠(Cynomys ludovicianus)和贝尔丁地鼠中,它们的母兽有共同的巢穴,并与自己的幼崽一起哺养与其无亲缘关系的幼崽。在阿根廷长耳豚鼠(Dolichotis patagonum)中,幼崽也被放置在共同的地道中,但母兽不允许除了自己的幼崽之外的其他幼崽哺乳。
物种保护
编辑物种现状
根据世界自然保护联盟(IUCN)数据,截至2022年全球范围内,啮齿目动物中的北松鼠(Sciurus vulgaris)、原仓鼠(Cricetus cricetus)、羽尾跳鼠(Stylodipus telum)等478个物种种群数量呈减少趋势,21个物种种群数量呈增加趋势,该目除此之外的动物类群数量较为稳定。
致危因素
已经灭绝的啮齿目有Oligoryzomys victus、Oryzomys antillarum和Rattus macleari等,主要是由于入侵一些如黑鼠、褐鼠或者獴等,使这些物种遭到灭绝。
而现存的啮齿目动物受到威胁的主要因素有:一是各类大鼠、小鼠和一些鸟类等被广泛用于研究,美国国家生物医学研究协会(NABR)估计,20世纪末的约23年间,研究使用了约1998万只大鼠和小鼠,占所有实验动物的95%;二是一些啮齿目动物被人为捕食而造成种群数量减少,如被捕食是温哥华岛野生温哥华岛旱獭(Marmota vancouverensis)数量下降的直接原因;三是栖息地被破坏使种群数量减少,如由于人类开发、城市发展以及转为农用和商用,大部分潮汐沼泽栖息地已经丧失,破坏了盐沼禾鼠(Reithrodontomys raviventris)的生存环境,使之数量下降严重。
保护等级
啮齿目动物中,2021年,河狸(Castor fiber)被中国《国家重点保护野生动物名录》列为国家一级保护野生动物,巨松鼠(Ratufa bicolor)列为国家二级保护野生动物。
截至2022年,啮齿目下有2375个种被列入《世界自然保护联盟(IUCN)濒危物种红色名录》ver3.1,其中1.6%为灭绝(EX)、2.5%为极危(CR)、6.1%为濒危(EN)、5.4%为易危(VU)、4.3%为近危(NT)、63.0%为无危(LC)、17.1%为数据缺乏(DD)。
2023年,啮齿目下46个种被列入中国《有重要生态、科学、社会价值的陆生野生动物名录》。
保护措施
一方面,啮齿目动物受到各国法律保护,如在美国,用于实验研究的各类大鼠和小鼠等啮齿目动物受到《动物福利法》(Animal Welfare Act)保护;如1984年,温哥华岛旱獭被《美国濒危物种法》列为濒危物种,2000年被加拿大濒危野生动物地位委员会(COSEWIC)列为濒危动物。
另一方面,一些啮齿目动物出现在其分布范围内的许多保护区中,如Sicista loriger存在于黑海生物圈保护区(Black Sea Biosphere Reserve)、卢甘斯克自然保护区(Lugansk Nature Reserve)、乌克兰草原保护区(Ukrainian Steppe Reserve)和第聂伯罗-奥廖尔自然保护区(Dnipro-Orel Nature Reserve)等保护区中,对其进行保护;水豚也存在于许多保护区中。
主要价值
编辑饰用价值
有少数种类的啮齿目动物毛皮可以利用,并作为其他食肉毛皮兽的食物,在自然界的生态系统中有重要的意义。如岩松鼠的冬毛是较好的毛皮,20世纪80年代时中国陕西省每年即可收购12万张左右。
竹鼠(Rhizomyidae)鼠皮张大,皮毛细软,光泽油润,色泽艳丽,底绒厚,皮板厚薄适中,易于鞣制;毛基为灰色,易于染色,是制裘衣、皮领、帽子的上等原料,尤其板皮是制上等皮革、鞋的好原料。其皮制成的夹克、长大衣,色泽光亮,平滑、轻软、耐磨,外观可与貂皮媲美。
食用价值
至少有89种啮齿目动物,主要是大囊啮齿类动物,如豚鼠、刺豚鼠(Dasyprocta Punctata)和水豚(Hydrochoerus hydrochaeris)等被人类食用。
豚鼠首次被人们驯养用作食物可以追溯到公元前约2500年,而到了公元前1500年,它已成为印加帝国的主要肉类来源。在秘鲁库斯科,豚鼠在烹饪中被广泛使用,如烤豚鼠。每年约有6400万只豚鼠被食用。
在古罗马时期,睡鼠被饲养在特殊的容器(gliraria)中,或者被放养在大型的户外围栏中,通过喂养核桃、栗子和橡子来使它们变得肥美而便于食用。在秋季,人们通常在睡鼠体脂最丰满的时候从野外捕捉它们,然后将它们烤熟,沾上蜂蜜,或者在烹饪时用猪肉、松子和其他调味料来填充它们。
在亚马逊流域等大型哺乳动物稀缺的地区,无尾刺豚鼠(Cuniculus paca)和刺豚鼠占土著居民年度狩猎捕捉的约40%,但在大型哺乳动物丰富的森林地区,这些啮齿目动物仅占年度捕捉的约3%。
实验价值
啮齿目动物广泛用作动物实验的模型生物。自从1828年首次使用白化突变的褐家鼠(Rattus norvegicus)进行研究以来,它们成为第一个纯粹为科学研究目的而被驯化的动物。家鼠是实验室中最常用的啮齿动物,全球估计每年使用五千万只家鼠进行科研。因其体型小巧、繁殖力强、怀孕周期短、易于操作,且容易受到许多与人类相关的疾病和感染的影响,家鼠在遗传学、发育生物学、细胞生物学、肿瘤学和免疫学等领域的研究中被广泛应用。
赏玩价值
啮齿目动物,包括豚鼠、南美洲栗鼠(Chin chilladale)、家鼠、仓鼠(Cricetinae)、沙鼠(Gerbillinae)、毛丝鼠(Chinchilla)、松鼠和花栗鼠(Tamias),因适合在狭小的空间中生活,且性情温顺,是便于饲养的宠物。
危害
编辑农林损失
一只大鼠每天可以吃掉大约100根4-6周龄的大米。啮齿目动物还对印度尼西亚的甘蔗种植园造成很大破坏,损失从2-100%不等。特别是如仓鼠(Cricetus cricetus)会破坏小麦和野豌豆、蚕豆、土豆、甜菜和其他作物的田地。在普通田鼠(Microtus arvalis)数量过剩的情况下,80%以上人工幼林可能会受到的损害,90%以上果树的有机种植园可能会受到损害。
除此之外,用于人类和动物食用的食物经常受到啮齿目动物的破坏或污染;该目动物由于需要不断磨尖门牙,其会损坏各种由天然和人造材料制成的物体(如木材、皮革、织物、塑料绝缘材料、尼龙袋、建筑材料、贵重珍贵的博物馆藏品、古董、图书馆书籍等);该目动物以植物的不同部分为食,从而破坏植物的种子、幼苗、树皮、根和芽。
疾病传播
啮齿目动物与人类疾病有密切关系。如鼠类是鼠疫、钩端螺旋体病、流行性出血热、蜱传斑疹热、狂犬病、鼠咬热、土拉伦斯病、布鲁氏杆菌病、恙虫病、地方性斑疹伤寒等的病原体的贮存宿主和传染源。
鼠疫
鼠疫(Plague)是由鼠疫杆菌(yersinia pestis)引起的一种烈性传染病,常流行于啮齿目动物,是一典型的自然疫源性疾病。在一定的地理景观中,病原体通过跳蚤在啮齿目动物间传播,致使出现地方性流行。啮齿目动物是形成鼠疫自然疫源性的重要环节,它必须与鼠疫杆菌、寄生蚤在特定的自然环境中同时并存,成为互相联系、彼此制约的统一体,才能使鼠疫病在某些地区长期保存下来和暴发流行。
危害
啮齿目动物是最主要的传染源。自然界对鼠疫易感的动物已知约有223种。一般分主要保菌鼠和次要保菌鼠两类。主要保菌鼠有黄鼠、长爪砂鼠(Meriones unguiculatus)、旱懒(Marmota)等,除易感性强外,还能常保持传播力较强的体外寄生蚤,因而可以保持疫源地的长期存在。次要保菌鼠有仓鼠、鼷鼠、白松鼠、小田鼠及家鼠等,虽易感,但不一定有传播力强的体外寄生蚤。肺鼠疫病人是最危险的传染源。腺鼠疫在未转入败血症型或肺型时,传染意义不大。潜伏期平均为3-5天,原发型肺鼠疫和原发型败血症鼠疫则仅有1天或更短。曾接种免疫者,潜伏期可延长9-12天。
由于病原体侵入机体的途径、数量、毒力及抗体抵抗等因素不同,鼠疫病原体在机体内形成鼠疫临床型的多样性。临床型最多见者为腺鼠疫,其次是肺鼠疫、败血型鼠疫,有时还可见到皮肤型鼠疫。鼠疫在人类历史上曾有多次严重流行,其中有三次是世界性大流行,死于鼠疫者近一亿数千万人,波及30多个国家。
类型
有腺型、肺型、败血型及轻型等,各型的初期症状相同,起病急,以畏寒发热开始,体温迅速上升达39-40℃。各型的特殊表现如下:
1、腺鼠疫:最常见。除全身中毒症状外,以急性淋巴结炎为特征,发生部位以腹股沟多见(70%)。淋巴结肿大在发病后第2-4天最为明显,与皮下及周围组织粘连,失去活动性,表面潮红,触之坚硬,有剧烈疼痛,患者往往处于强迫体位,为腺鼠疫的特征。如治疗不及时,结肿化脓、破溃,病情逐渐加重,可因严重毒血症与心功能不全而死亡。
2、肺鼠疫:原发性肺鼠疫是鼠疫中最严重的一型,病死率可高达70-100%,流行病学方面的危害性最大。除全身中毒症状外,有剧烈胸痛、咳嗽、咳痰,并很快有大量泡沫样血痰,内含有大量鼠疫杆菌。患者呼吸极为困难,颜面及全身皮肤发继迅速加重,死后皮肤常呈黑紫色,故有“黑死病”之称。继发性肺鼠疫多由腺鼠疫演变而来,故兼有腺鼠疫及肺鼠疫的症状,为造成原发性肺鼠疫流行的基本传染源。
3、败血型鼠疫:最凶险的一型,被称为“暴发型鼠疫”。由于细菌血源性扩散,多个器官被感染,特别是脾脏和肺部,患者可能出现败血性休克,表现为发热、厌食、呕吐、腹泻、心跳加速、脉搏减弱、低血压、末梢肢体发冷和白细胞数量明显增多等典型败血症症状。患者通常在出现细菌血症后1到2天内死亡。
4、其它如皮肤型、脑膜型、肠型、眼型等均少见。
防治
人对鼠疫普遍易感。经接种鼠疫菌苗后,可以产生一定时期的免疫力。鼠疫活菌苗对腺鼠疫可降低易感性,对肺鼠疫不能预防。患者应隔离治疗。除此之外,也可用药物防治。
流行性出血热
流行性出血热(Epidemic Hemorrhagic Fever,EHF)1932年在中国黑龙江流域和苏联远东地区发现,曾称为“远东出血性肾病肾炎”。1942年定名为“流行性出血热”。以下简称出血热。
危害
小型啮齿目动物黑线姬鼠是本病的主要传染源,但本病可能为多宿主性。急性病人的血及尿污染医护人员的伤口而感染本病,虽有报告但非自然传播,故人作为传染源的可能性很小。除野鼠外,家鼠亦是本病的宿主动物。不同的性别、年龄、职业的人均可以感染本病,二次发病罕见。病后有巩固免疫力。在老疫区居民发病比新迁入的人发病少,说明本病有隐性感染存在。疫源地的特点是地势低注、潮湿、多水、多草或存在成片的荒草地带。全年各月均有病例发生。发病高峰可因地而异。发病通常以青壮年为主,多数为野外工作的劳动者,如水利建设、农垦劳动、开发森林、开垦荒地、野营部队等均易与鼠类接触,一般是男性多于女性。
潜伏期5-46天,一般为7-14天,以2周为最多见。主要表现为先有3-7天短期的发热和中毒症状,以后出现低血压休克和肾功能损害。本病过程分五期:即发热期、低血压休克期、少尿期、多尿期和恢复期。由于病变轻重不同,故各期轻重也有差异。
防治
主要应做好灭鼠防鼠、疫情(人间、鼠间)监测、灭螨防螨、个人卫生和个人防护,加强消毒(病人的血尿、死鼠、宿主动物的排泄物)野外作业人员和工地的预防措施。本病无特异性病原疗法,主要进行综合性、预防性治疗。
代表物种
编辑啮齿目动物数量较多,故仅选取以下几种研究较多的常见物种进行介绍:红腿长吻松鼠(Dremomys pyrrhomerus)、倭松鼠(Tamiops maritimus)、豪猪(Hystrix hodgsoni)、中华竹鼠(Rhizomys sinensis)、大足鼠(Rattus nitidus )、河狸(Castor fiber)。
红腿长吻松鼠
红腿长吻松鼠为松鼠科(Sciuridae)长吻松鼠属(Dremomys),体重为240-295克,体长194-215毫米,尾长138-152毫米。吻细长,似锥形,背中央颜色较深,腹部淡黄白色。其为中国特有种,主要栖居在密林中,多喜晨、昏活动,半树栖种类,在树洞或石隙中筑巢,冬季活动较少。每年换毛2次。主要以摄食各种坚果及浆果为主,亦食各种树叶、嫩枝、花芽及鸟卵、雏鸟和昆虫等。每年繁殖2次,每次产3-4只仔,以2仔居多。

豪猪
豪猪为豪猪科(Hystricidae)豪猪属(Hystrix)动物,头小,眼小,耳廓短,体形小,身体长,四肢粗壮;背部与尾部生有长而硬的棘刺,尾末端覆有白色棘簇,棘的后部具有很多“珠串”状的球节;背面的棘刺比较扁,上面有沟;腹部的棘刺柔软纤细。其广布于中国南部和中部,包括海南岛,延伸到尼泊尔、印度、缅甸、泰国、马来西亚、苏门答腊和婆罗洲。栖息在森林和开阔田野。在堤岸和岩石下挖洞,穴居。晚上出洞,沿着固定的路线觅食。豪猪不会扇动它们的棘刺,但遇危险时以后退方式御敌。报警时摇动尾棘作响,喷鼻息和踩脚。

倭松鼠
倭松鼠是松鼠科(Sciuridae)花松鼠属(Tamiops)动物,体长115-158mm,尾长85-130mm。全身自额部起,向后至尾部呈灰褐色,并杂有黑毛。背部正中有一条明显的黑色条纹,两侧有两条褐黄或浅黄的纵纹,次外侧有两条黑褐色纵纹,最外侧为两条浅黄或淡黄白色纵纹。其分布于中国南部,延伸至缅甸、越南和老挝。生活在次生林带的常绿阔叶林和混交林中,有时食植物的花蜜。高度树栖性,但常在地面活动,通常呈跳跃式活动。在攀登树干时,常以“之”字形沿树干向上爬。下树时,头朝下,迅速地移到树干基部,最后跳到地面活动。1年繁殖2次,春季和秋季各1次。

中华竹鼠
中华竹鼠是竹鼠科(Rhizomyidae)竹鼠属(Rhizomys)动物,头体长216-380毫米,尾长50-96毫米,体重1.875-1.95千克。额和面侧颜色较深。耳短,突出于毛外。腹毛稀少。皮毛柔软,背部和体侧浅棕灰色,尾短而裸露。其在世界范围内分布于缅甸北部、越南和中国等。中国分布于南部和西南部,通常生活在高海拔竹林,也可生活在松林。主要食物是竹根和竹笋。大多数在地面以上觅食。在松软的土壤上挖洞,单独居住(交配时例外)。窝内垫以竹叶,所有的洞系都有一个逃生通道。大多数洞约用1年,此后居住者因为食物资源的消耗而转移到一个新的地方。每胎最多达8只,繁殖高峰在春季。幼仔出生时无毛,3月龄断奶。

大足鼠
大足鼠是鼠科(Muridae)家鼠属(Rattus)动物,外形体型中等,形似褐家鼠(Rattus norvegicus),耳背毛色深暗。雌鼠乳头6对。毛色背部毛色暗棕褐色,略带黄色色调,具棕褐色和黑色长毛。尾较长,几接近体长,单色,背面毛色稍暗。世界范围内分布于尼泊尔、印度、缅甸、泰国和中国。中国主要见于长江以南广大地区,栖息于海拔800-3200米的山地、丘陵地带环境中。营巢于田便、水沟旁、溪流附近及草垛下。常在玉米、豆类等农田中觅食,对农作物危害很严重。主要以种子为食,喜食玉米和稻谷,亦食浆果、草籽和其他小型鼠类及鱼、螃蟹等。大足鼠主要在夜间活动,白天偶尔可见。4、5月为繁殖高峰期,平均每胎产仔4-6只,最多为12只。

河狸
河狸是河狸科(Castoridae)河狸属(Castor)动物,体长0.6-1米,体重17-30千克,体型肥壮,第四趾十分特殊,有双爪甲,一为爪形,一为甲形。尾大而宽,上下扁平,上覆大型角质鳞片和稀疏的毛。体背毛色由土黄、棕色到暗褐色。世界范围内分布于俄罗斯、德国、法国、芬兰、波兰、加拿大和中国等。中国分布于新疆、内蒙古等地。栖息在寒温带针叶林和针阔混交林林缘的河边。岸边筑洞,夜间活动,营半水栖生活,善游泳和潜水,不冬眠,自卫能力很弱。寿命为12-20年。

类群对比
编辑树鼩目(Scandentia)和翼手目(Chiroptera)均为哺乳纲(Mammalia)下目。其中,树鼩目的形态与习性与啮齿目的松鼠相似,而翼手目动物数量较多,是哺乳动物中数量仅次于啮齿目的第二大类群,对其进行对比如下:
名称 | 形态特征 | 分布栖息 | 生活习性 | 图片 |
啮齿目(Rodentia) | 啮齿目动物通常很小,乳头2-10对,以4对居多。前肢短,5指(趾)居多,指(趾)端具爪。被毛结构和毛色变化复杂而多样 | 分布于除南极洲、新西兰和一些海洋岛屿以外的世界各地 | 性怯弱,行动敏捷,有些种类营日间活动,有些种类只在夜间活动 | 花栗鼠(Tamias striatus)
|
翼手目(Chiroptera) | 前肢特化,尤其是骨骼有较大的变化。牙齿齿冠原始,白齿齿尖和齿崎连接成“W”形。胸部1对乳头。耳壳发达,常具发达的耳屏或对耳屏 | 除两极外,东西两半球都有分布。大多数种类见于热带和温带 | 唯一真正具有飞行能力的哺乳动物夜行性,能发出和回收超声波以辨别方向,是适应性最强的哺乳动物之一 | Corynorhinus townsendii
|
树鼩目(Scandentia) | 吻部狭窄,脑颅宽大,上门齿犬齿状,而犬齿缩小;下白齿保留基本的食虫模式。四肢五趾,趾有强而弯曲的爪而无甲,后肢长 | 主要分布于亚洲东南部森林中 | 在树上和地上觅食。性贪食,食性广泛,尤其喜食动物性食物和果实。昼行性,鸣叫声特高 | 北树鼩(Tupaia belangeri)
|



参考资料
编辑展开[1]Rodentia Bowdich, 1821.itis. [2023-09-19].
[2]Delaney A, Treuting M, Rothenburger L. Pathology of Wildlife and Zoo Animals[M]. 2018: 499-513. [2023-09-19].
[3]Rodentia.Animal Diversity Web. [2023-09-19].
[4]Rodentia - OrderRemove.iucnredlist. [2023-09-19].
[5]国家重点保护野生动物名录.国家林业和草原局 国家公园管理局. [2023-09-19].
[6]路纪琪, 吕国强, 李新民主编. 河南啮齿动物志[M]. 郑州: 河南科学技术出版社, 1997.10: 96-97. (2)
[7]罗蓉等编著. 贵州兽类志[M]. 贵阳: 贵州科技出版社, 1993.10: 185.
[8]吴跃峰, 武明录, 曹平萍等编著. 河北动物志 两栖、爬行、哺乳动物类[M]. 石家庄: 河北科技出版社, 2009.11: 186.
[9]全理华, 张兴旺等. 东北地区主要医学动物与疾病[M]. 沈阳: 白山出版社, 1990.05: 404-409+413-418. (9)
[10]Jurišić A, Ćupina A I, Kavran M, et al. Surveillance Strategies of Rodents in Agroecosystems, Forestry and Urban Environments[J]. Sustainability, 2022, 14(15): 9233. [2023-09-19].
[11]Jerry O, WOLFF. Social biology of rodents[J]. Integrative Zoology, 2007, 2(4): 193-204. [2023-09-19].
[12]董yu茂主编, 毛节荣等编. 浙江动物志 兽类[M]. 杭州: 浙江科学技术出版社, 1989.11: 63-64.
[13]Kay E H, Hoekstra H E. Rodents[J]. Current Biology, 2008, 18(10): R406-R410. [2023-11-02].
[14]啮齿目(化石).中国大百科全书. [2023-09-19].
[15]Wood D J. The extinction of the Multituberculates outside North America: a global approach to testing the competition model[D]. The Ohio State University, 2010[2023-11-02].
[16]Asher R J, Meng J, Wible J R, et al. Stem Lagomorpha and the antiquity of Glires[J]. Science, 2005, 307(5712): 1091-1094. [2023-11-02].
[17]Samuels J X, Zancanella J. An early Hemphillian occurrence of Castor (Castoridae) from the Rattlesnake Formation of Oregon[J]. Journal of Paleontology, 2011, 85(5): 930-935. [2023-11-02].
[18]Marivaux L, Marzougui W, Ammar H K, et al. A new and primitive species of Protophiomys (Rodentia, Hystricognathi) from the late middle Eocene of Djebel el Kébar, Central Tunisia[J]. Palaeovertebrata, 2014, 38(1): e2. [2023-11-02].
[19]Gheerbrant E, Rage J C. Paleobiogeography of Africa: how distinct from Gondwana and Laurasia?[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2006, 241(2): 224-246. [2023-11-02].
[20]Vélez-Juarbe J, Martin T, Macphee R D E, et al. The earliest Caribbean rodents: Oligocene caviomorphs from Puerto Rico[J]. Journal of Vertebrate Paleontology, 2014, 34(1): 157-163. [2023-11-02].
[21]Schenk J J, Rowe K C, Steppan S J. Ecological opportunity and incumbency in the diversification of repeated continental colonizations by muroid rodents[J]. Systematic biology, 2013, 62(6): 837-864. [2023-11-02].
[22]Rodentia.tolweb. [2023-11-02].
[23]Entries linking to rodent.etymonline. [2023-11-02].
[24]罗建南, 邓学建. 南山国家公园脊椎动物资源及多样性现状[M]. 长沙: 湖南科学技术出版社, 2020.08: 175.
[25]郑斌, 赵欣如, 宋大昭主编. 小五台山陆生脊椎动物资源调查[M]. 石家庄: 河北科学技术出版社, 2016.05: 63-67. (6)
[26]Landry Jr S O. The Rodentia as omnivores[J]. The Quarterly Review of Biology, 1970, 45(4): 351-372. [2023-11-02].
[27]NORTHERN GRASSHOPPER MOUSE.Natural Science Research Laboratory. [2023-11-02].
[28]Jarvis J U M. Eusociality in a mammal: cooperative breeding in naked mole-rat colonies[J]. Science, 1981, 212(4494): 571-573. [2023-11-02].
[29]Hoogland J L. Sexual dimorphism of prairie dogs[J]. Journal of Mammalogy, 2003, 84(4): 1254-1266. [2023-11-02].
[30]Vaughan T A. Reproduction in the plains pocket gopher in Colorado[J]. Journal of Mammalogy, 1962, 43(1): 1-13. [2023-11-02].
[31]Holmes W G, Mateo J M. Kin recognition in rodents: issues and evidence[J]. Rodent societies, 2007: 216-228. [2023-11-02].
[32]Vaché M, Ferron J, Gouat P. The ability of red squirrels (Tamiasciurus hudsonicus) to discriminate conspecific olfactory signatures[J]. Canadian Journal of Zoology, 2001, 79(7): 1296-1300. [2023-11-02].
[33]Shelley E L, Blumstein D T. The evolution of vocal alarm communication in rodents[J]. Behavioral Ecology, 2005, 16(1): 169-177. [2023-11-02].
[34]In Altenmüller, Eckart, Schmidt, Sabine, Zimmermann, Elke (eds. The Evolution of Emotional Communication: From Sounds in Nonhuman Mammals to Speech and Music in Man[M]. The Evolution of Emotional Communication, 2013: 123-124. (2)
[35]Vanden Hole C, Van Daele P A A G, Desmet N, et al. Does sociality imply a complex vocal communication system? A case study for Fukomys micklemi (Bathyergidae, Rodentia)[J]. Bioacoustics, 2014, 23(2): 143-160. [2023-11-02].
[36]Long C V. Vocalisations of the degu Octodon degus, a social caviomorph rodent[J]. Bioacoustics, 2007, 16(3): 223-244. [2023-11-02].
[37]Ancillotto L, Sozio G, Mortelliti A, et al. Ultrasonic communication in Gliridae (Rodentia): the hazel dormouse (Muscardinus avellanarius) as a case study[J]. Bioacoustics, 2014, 23(2): 129-141. [2023-11-02].
[38]Haverkamp S, Wässle H, Duebel J, et al. The primordial, blue-cone color system of the mouse retina[J]. Journal of Neuroscience, 2005, 25(22): 5438-5445. [2023-11-02].
[39]What Do Rats See?.ratbehavior. [2023-11-02].
[40]Vibrational communication in mammals.mapoflife. [2023-11-02].
[41]Randall J A. Evolution and function of drumming as communication in mammals[J]. American Zoologist, 2001, 41(5): 1143-1156. [2023-11-02].
[42]In MacDonald, D. The Encyclopedia of Mammals[M]. 2001: 578-587. (10)
[43]Waterman J, Wolff J O, Sherman P W. Male mating strategies in rodents[J]. Rodent societies, 2007: 27-41. [2023-11-02].
[44]McGuire B, Bemis W E. Parental care[J]. Rodent societies, 2007: 231-242. [2023-11-02].
[45]Gilbert A N. Mammary number and litter size in Rodentia: the “one-half rule”[J]. Proceedings of the National Academy of Sciences, 1986, 83(13): 4828-4830. [2023-10-18].
[46]罗建南邓学建. 南山国家公园脊椎动物图鉴[M]. 长沙: 湖南科学技术出版社, 2020.08: 166-169.
[47]王岐山主编. 安徽兽类志[M]. 合肥: 安徽科学技术出版社, 1990.03: 175-178. (3)
[48]万冬梅主编. 国家生态保护丛书 国家保护动物卷[M]. 2015.12: 76-77. (2)
[49]In Wolff, Jerry O. Rodent Societies: An Ecological and Evolutionary Perspective[M]. Rodent Societies: An Ecological and Evolutionary Perspective[M], 2007: 3-8,226–227. (8)
[50]Oligoryzomys victus.IUCN. [2023-10-18].
[51]Oryzomys antillarum.IUCN. [2023-10-18].
[52]Rattus macleari.IUCN. [2023-10-18].
[53]Frankie L, Barbara A. Regulation of Rodents[J]. Science, 1999: 1463-1463. [2023-09-19].
[54]Marmota vancouverensis.iucnredlist. [2023-09-19].
[55]Reithrodontomys raviventris.iucnredlist. [2023-09-19].
[56]有重要生态、科学、社会价值的陆生野生动物名录.国家林业和草原局 国家公园管理局. [2023-09-19].
[57]Sicista loriger.IUCN. [2023-10-18].
[58]Hydrochoerus hydrochaeris.iucnredlist. [2023-09-19].
[59]陕西省动物研究所主编. 陕西珍贵经济兽类图志[M]. 西安: 陕西科学技术出版社, 1981.11: 13.
[60]宋兴超, 杨福合, 邢秀梅, 岳志刚. 我国竹鼠资源种类、价值及人工驯养前景[J]. 特种经济动植物, 2009, 12(02): 8-10. [2023-10-18].
[61]Fiedler L A. Rodents as a food source[C]. /Proceedings of the Vertebrate Pest Conference, 1990[2023-11-02].
[62]Hedrich H J. History, strains and models[M]. 2000: 3-16. [2023-11-02].
[63]In Foster, Henry. The Mouse in Biomedical Research: History, Genetics, and Wild Mice[M]. The Mouse in Biomedical Research: History, 1981: 1-5. (5)
[64]Pet Rodents.RSPCA. [2023-11-02].
[65]Guinea pigs.RSPCA. [2023-11-02].
[66]王海洋、姜建波、李春花. 小动物疾病学[M]. 武汉: 华中科技大学出版社, 2020: 49.
[67]汪春晖, 张锦海, 叶福强主编. 传染病诊疗与社区防控指南[M]. 苏州: 苏州大学出版社, 2020: 62-68. (7)
[68]王廷正, 许文贤主编. 陕西啮齿动物志[M]. 西安: 陕西师范大学出版社, 1993.04: 178-180. (2)
[69]红腿长吻松鼠(Dremomys pyrrhomerus).inaturalist. [2023-09-19].
[70]大足鼠(Rattus nitidus).inaturalist. [2023-09-19].
[71]河狸(Castor fiber).inaturalist. [2023-09-19].
[72]树鼩目(Scandentia).ITIS. [2023-10-18].
[73]翼手目(Chiroptera).ITIS. [2023-10-18].
[74]盛和林, 王培潮等. 哺乳动物学概论[M]. 上海: 华东师范大学出版社, 1985.06: 70-71. (2)
内容由百科小编提供,本内容不代表globalbaike.com立场,内容投诉举报请联系全球百科客服。如若转载,请注明出处:https://globalbaike.com/26665/